Катаболизм (расщепление) жирных кислот

Образовавшийся ацетил-КоА включается в цикл трикарбоновых кислот. Укороченный на два углеродных атома ацил-КоА снова проходит весь путь Р-окисления вплоть до образования 4-углеродного соединения бутирил-КоА. Бутирил-КоА затем окисляется до 2 молекул ацетил-КоА, которые также включаются в цикл трикарбоновых кислот. На этой стадии катаболизм жирной кислоты завершается. Окисление ненасыщенных жирных… Читать ещё >
Катаболизм (расщепление) жирных кислот (реферат, курсовая, диплом, контрольная)
Катаболизм высших жирных кислот осуществляется преимущественно путем окисления. Ненасыщенные высшие жирные кислоты предварительно восстанавливаются до предельных кислот. Предельные (насыщенные) жирные кислоты окисляются ступенчато, путем отщепления от их молекул ацильных 2-углеродных фрагментов СН3-СО- (рис. 9.15). Распад высших жирных кислот проходит в несколько стадий. Все стадии окисления ускоряются специфическими ферментами.
Начальная стадия катаболизма жирных кислот заключается в активации. Свободная жирная кислота независимо от длины углеводородной цепи химически инертна и не может подвергаться биохимическим превращениям, пока не будет активирована. Активация жирной кислоты R-COOH протекает на наружной поверхности мембраны митохондрий при участии АТР, кофермента-А (HS-KoA) и ионов Mg2'. Реакция катализируется ферментом ацил-КоА-синтетазой:
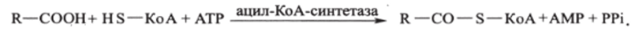
В результате образуется ацил-КоА (R-CO-S-KoA) — активная коэнзимная форма кислоты.
Дальнейшее окисление жирных кислот происходит на внутренней стороне мембраны митохондрий.
Коэнзимные формы кислот ацил-КоА, как и свободные жирные кислоты, не обладают способностью проникать внутрь митохондрий. Из цитоплазмы к внутренней стороне мембраны митохондрий ацил-КоА переносится карнитином. Вначале карнитин реагирует с ацил-КоА:
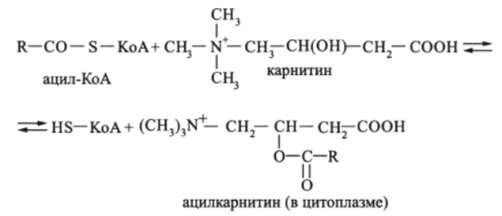
В результате в цитоплазме образуется ацилкарнитин. который проходит через мембрану внутрь митохондрии, где распадается, высвобождая ацил-КоА:
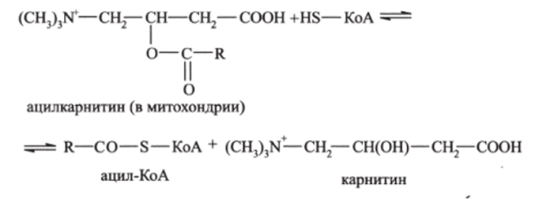
Окисление жирных кислот внутри митохондрий протекает через несколько последовательных стадий.
Первая стадия. Ацил-КоА реагирует с FAD и подвергается ферментативному отщеплению водорода — дегидрированию. При этом ацил-КоА теряет 2 атома водорода (окисляется), превращаясь в КоА-эфир ненасыщенной кислоты еноил-КоА, соединение с двойной связью:
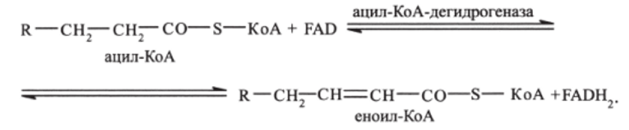
Следует отмстить, что существует несколько ацил-КоА-дегидрогеназ, каждая из которых обладает специфичностью по отношению к ацил-КоА с определенной длиной углеродной цепи.
Вторая стадия. Еноил-КоА (при участии фермента еноил-КоА-гидратазы) присоединяет молекулу воды. Происходит гидратация двойной связи. В результате образуется р-оксиацил-КоА:
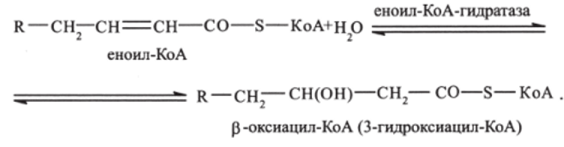
Третья стадия. р-оксиацил-КоА дегидрируется с образованием 3-оксиацилКоА (катализируется ферментом N, А Независимой дегидрогеназой):
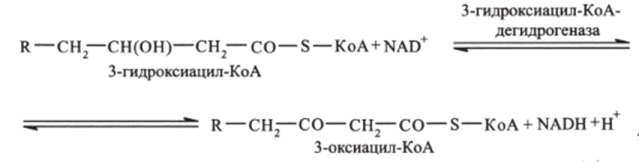
Четвертая стадия — тиолазная реакция. В ходе тиолазной реакции происходит расщепление 3-оксиацил-КоА с образованием укороченной на два углеродных атома молекулы ацил-КоА и ацильного 2-углеродного фрагмента СН3—СО-, связанного в ацетил-КоА (реакция катализируется ацетил-КоА-ацилтрансферазой):
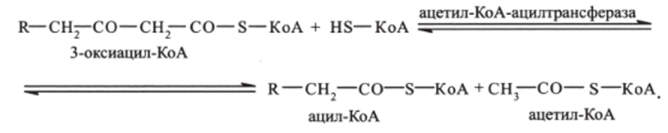
Образовавшийся ацетил-КоА включается в цикл трикарбоновых кислот. Укороченный на два углеродных атома ацил-КоА снова проходит весь путь Р-окисления вплоть до образования 4-углеродного соединения бутирил-КоА. Бутирил-КоА затем окисляется до 2 молекул ацетил-КоА, которые также включаются в цикл трикарбоновых кислот. На этой стадии катаболизм жирной кислоты завершается.
В каждом цикле расщепления жирных кислот образуется р-кетоацил-КоА со все более укороченной углеводородной цепью, поэтому рассмотренный процесс в целом можно называть Р-окислением жирных кислот. NADH и FADH2, образующиеся при Р-окислении, затем отдают свои электроны митохондриальной дыхательной цепи.
При окислении жирной кислоты, содержащей п углеродных атомов, происходит (л/2−1) цикл Р-окисления. Например, при окислении пальмитиновой кислоты (С|6) повторяется 7 циклов р-окисления. Следовательно, суммарное уравнение р-окисления активированной пальмитиновой кислоты имеет вид.
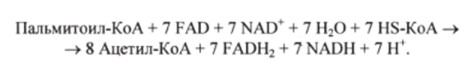
Молекула пальмитиновой кислоты превращается в 8 молекул ацетил-КоА и дополнительно 7 молекул NADH и 7 молекул FADH2. Затем при окислении NADH синтезируются 2,5 молекулы АТР, а при окислении FADH2 — 1,5 молекулы АТР. Таким образом, окисление 1 молекулы пальмитиновой кислоты приводит к синтезу 106 молекул АТР из ADP и Р, (с учетом затраты 2 молекул АТР на образование 1 молекулы пальмитоил-КоА и синтеза 1 молекулы GTP на каждую молекулу ацетил-КоА в цикле лимонной кислоты). Это означает, что энергетический выход биоокисления пальмитиновой кислоты составляет 33%.
Следует отметить, что если окончательным продуктом р-окисления высших жирных кислот с четным числом углеродных атомов является ацетил-КоА, то с нечетным — пропионил-КоА.
Окисление ненасыщенных жирных кислот, в принципе, происходит так же, как и окисление насыщенных, но имеет ряд особенностей. До двойной связи цепь такой кислоты укорачивается в результате обычного р-окисления с образованием /*мс-еноил-КоА. Далее фермент изомераза отодвигает двойную связь и образует трансизомер активированной кислоты т/?анс-еноил-КоА:
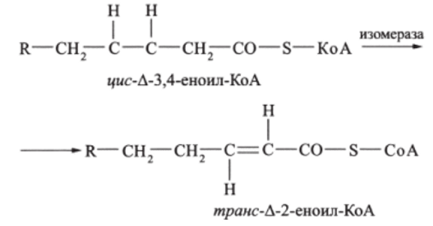
При р-окислении жирных кислот, имеющих две и более ненасыщенных связи, требуется еще один дополнительный фермент.
Природные масла содержат главным образом жирные кислоты с четным числом углеродных атомов. Однако в маслах многих растений и некоторых морских организмов присутствуют жирные кислоты с нечетным числом атомов углерода. Кроме того, у жвачных животных при переваривании углеводов в рубце образуется большое количество пропионовой кислоты СН3СН2СООН с тремя углеродными атомами. Пропионат всасывается в кровь и окисляется в печени и других тканях.
Жирные кислоты с нечетным числом углеродных атомов окисляются таким же образом, как и жирные кислоты с четным числом, с той лишь разницей, что на последнем этапе расщепления (р-окисления) образуется одна молекула пропионилКоА и одна молекула ацетил-КоА, а не две молекулы ацетил-КоА.
Активированный 3-углеродный фрагмент — пропионил-КоА — включается в цикл трикарбоновых кислот после превращения в сукцинил-КоА.
В реакции изомеризации метилмалонил-КоА участвует кофермент дезоксиаденозилкобаламин, производное витамина В12.
Пропионат образуется также при расщеплении четырех аминокислот — валина, изолейцина, метионина, треонина — и из холестерина.