Эритроциты.
Физиология человека и животных

Эритропоэз значительно возрастает после кровопотери, при патологическом укорочении жизни эритроцитов, а также при уменьшении количества кислорода, поступающего с вдыхаемым воздухом (например, в условиях высокогорья, когда усиленный эритропоэз является важным фактором адаптации организма человека). Стимулом к ускоренной дифференцировке и размножению предшественников эритроцитов в костном мозге… Читать ещё >
Эритроциты. Физиология человека и животных (реферат, курсовая, диплом, контрольная)
Эритроциты являются самыми многочисленными и высокоспециализированными клеточными элементами крови. У взрослых мужчин в 1 мкл крови содержится в среднем 5,1 млн эритроцитов, а у женщин — 4,6 млн. Основная часть эритроцитов (85%) имеет форму двояковогнутых дисков. Такие клетки называются нормоцитами. Их диаметр составляет 7—8 мкм, толщина в центральной области — около 2 мкм (рис. 4.2, а). Оставшиеся 15% эритроцитов могут иметь увеличенный или уменьшенный размер, круглую форму (сфероциты), отростки и др.
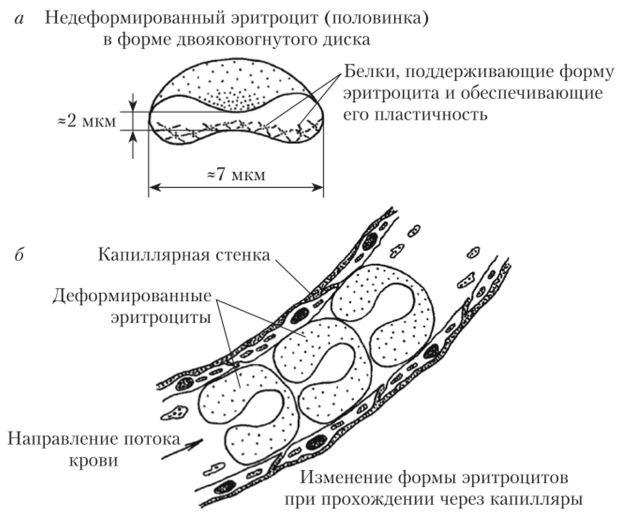
Рис. 4.2. Размеры и форма эритроцитов (а), изменение формы эритроцитов при прохождении через капилляры (б).
Благодаря своему строению нормоцигы оптимально приспособлены к выполнению газотранспортной функции. Поверхность двояковогнутого диска больше, чем поверхность шара такого же объема, а расстояние, которое должны пройти газы от наружной мембраны эритроцита до транспортных белков, — меньше, чем у клеток шаровидной формы. Таким образом, диффузионная поверхность увеличивается, а диффузионное расстояние — уменьшается. Суммарная площадь поверхности всех эритроцитов крови взрослого человека составляет около 3800 м2.
Форма эритроцитов и особенности строения мембран обеспечивают им такое свойство, как пластичность, т.е. способность к обратимой деформации (изменению формы). Это позволяет эритроцитам успешно проходить по капиллярам, имеющим значительно меньший диаметр (до 2—3 мкм)
и изогнутую форму (рис. 4.2, б). Пластичность эритроцитов связана с присутствием в их мембране белков гликофоринов, а в цитоплазме — белков спектринов, которые тесно взаимодействуют друг с другом. В процессе старения клеток происходит связывание спектрина с гемоглобином цитоплазмы. Это затрудняет нормальное взаимодействие спектрина и гликофорина, вызывает нарушение формы и пластичности эритроцитов. Из нормоцитов они превращаются в сфероциты, что является одной из причин задержки и разрушения таких клеток в селезенке. Помимо этого, гликофорины мембран эритроцитов придают их наружным новерностям гидрофильные свойства и создают на них небольшой отрицательный электрический потенциал ((^-потенциал; «дзета"-потенциал). Благодаря этому эритроциты отталкиваются друг от друга, образуя в плазме устойчивую суспензию.
Уникальность эритроцитов как клеток состоит в том, что в зрелом состоянии они не имеют ядра и цитоплазматических органелл. По этой причине эритроциты не способны к синтезу белков или липидов, а также аэробному окислительному фосфорилированию. Главным источником энергии в зрелых эритроцитах, как и в других клетках, является АТФ. Это вещество необходимо им, в частности, для активного транспорта ионов через мембрану. АТФ в эритроцитах образуется за счет анаэробного (без участия кислорода) окисления глюкозы (гликолиза). Это резко уменьшает собственные потребности красных клеток крови в кислороде, которые составляют не более 2% от всего транспортируемого газа. Другим следствием отсутствия у зрелых эритроцитов ядра и клеточных органелл является возможность размещения дополнительного количества гемоглобина. В зрелом эритроците масса гемоглобина составляет около 98% от массы всех белков цитоплазмы.
Основными функциями эритроцитов являются транспортная, гомеостатическая и защитная. Транспортная функция заключается в способности переносить кислород и углекислый газ, питательные вещества (белки и углеводы), биологически активные вещества. Гомеостатическая функция эритроцитов обусловлена тем, что содержащийся в них гемоглобин может связывать углекислый газ, снижая тем самым концентрацию в крови угольной кислоты и стабилизируя pH. Защитная функция эритроцитов состоит в их участии в процессах свертывания крови, в способности связывать и обезвреживать некоторые токсины, а также в наличии на мембранах специфических антигенов, которые делают эритроциты участниками ряда иммунологических реакций.
Образование эритроцитов (эритропоэз) происходит в кроветворных (гемопоэтических) тканях: желточном мешке у эмбриона, печени и селезенке у плода и в красном костном мозге у ребенка и взрослого человека. Во всех этих органах содержатся полипотентные стволовые гемопоэтические клетки — общие предшественники всех клеток крови. Стволовые клетки производят эритробласты — специализированные клетки, из которых могут развиваться уже только эритроциты (рис. 4.3).
Эритробласты через ряд промежуточных этапов роста и развития образуют безъядерные ретикулоциты, которые выходят в кровь и превращаются через 24—36 ч в зрелые эритроциты. Срок жизни созревших эритроцитов — 3—4 мес., после чего они захватываются и разрушаются клетками фагоцитирующей системы селезенки, красного костного мозга и печени. Способностью разрушать эритроциты обладают не только эти органы, но и большинство тканей организма, что наглядно проявляется при постепенном исчезновении подкожных кровоизлияний («синяков»). Интенсивность эритропоэза и разрушения эритроцитов очень велика: за одну минуту в организме взрослого человека образуется 160 * 106 эритроцитов, а за сутки обновляется около 0,8—1% от их общего количества, которое составляет приблизительно 25 • 1012 клеток.
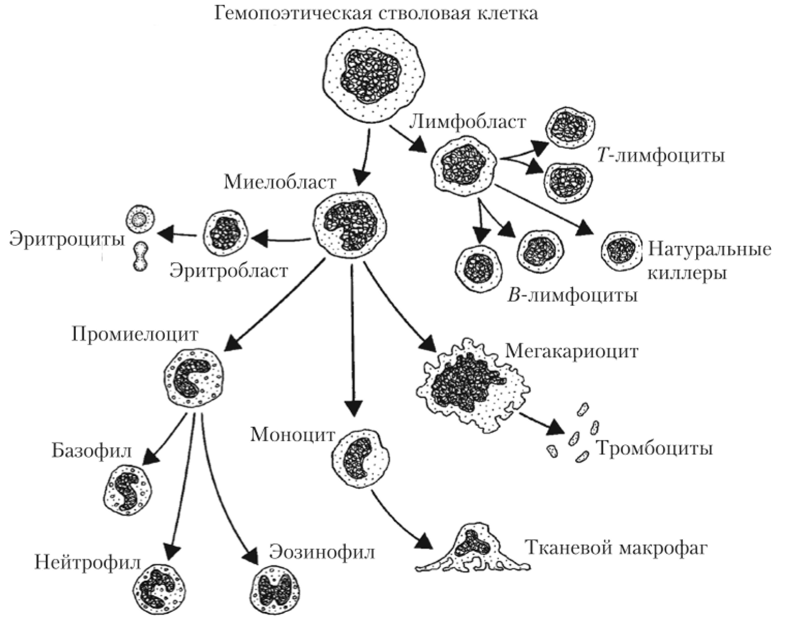
Рис. 43. Схема образования форменных элементов крови из стволовой гемопоэтической клетки.
Эритропоэз значительно возрастает после кровопотери, при патологическом укорочении жизни эритроцитов, а также при уменьшении количества кислорода, поступающего с вдыхаемым воздухом (например, в условиях высокогорья, когда усиленный эритропоэз является важным фактором адаптации организма человека). Стимулом к ускоренной дифференцировке и размножению предшественников эритроцитов в костном мозге является гликопротеин эритропоэтин. Главную роль в синтезе эритропоэтина играют почки, но в небольших количествах он образуется и в печени. Действие эритропоэтина усиливается многими «классическими» гормонами, например андрогенами, тироксином, гормоном роста. Различие в числе эритроцитов в крови мужчин и женщин обусловлено тем, что андрогены усиливают синтез эритропоэтина, а эстрогены его тормозят.
В последние годы эритропоэтин получил известность как один из запрещенных видов спортивного допинга. Его введение в организм спортсмена в период тренировок постепенно увеличивает количество эритроцитов в крови. К моменту соревнований такой спортсмен получает преимущество, лучше выполняя физическую работу в условиях ограниченного дыханием поступления кислорода. На сходном принципе основан «кровяной допинг». У спортсмена заранее несколько раз берется кровь и создается банк эритроцитов. Перед соревнованиями эти эритроциты вводятся ему в кровь, что повышает ее кислородную емкость и дает дополнительное преимущество.
Гемоглобин и его соединения, сконцентрированные в цитоплазме эритроцитов, обеспечивают их жизненно важные функции — дыхательную и гомеостатическую. Гемоглобин может успешно работать, только находясь в эритроцитах. В случае разрушения эритроцитов (гемолиза) и появления гемоглобина в плазме он очень быстро связывается с белком гаптоглобином, образуя комплекс, который нейтрализуется клетками фагоцитирующей системы печени и селезенки. Период полувыведения гемоглобина из кровотока составляет около 10 мин. Локализация гемоглобина внутри эритроцита позволяет защитить его от воздействия активных компонентов плазмы, сконцентрировать и создать оптимальные условия для функционирования. Кроме того, гемоглобин сложным образом взаимодействует с мембраной эритроцита и элементами цитоскелета, что способствует повышению его активности.
Молекула гемоглобина состоит из белковой части (глобина), которая включает две пары иолииеитидных цепей, и небелковой части, состоящей из четырех гемов (рис. 4.4).
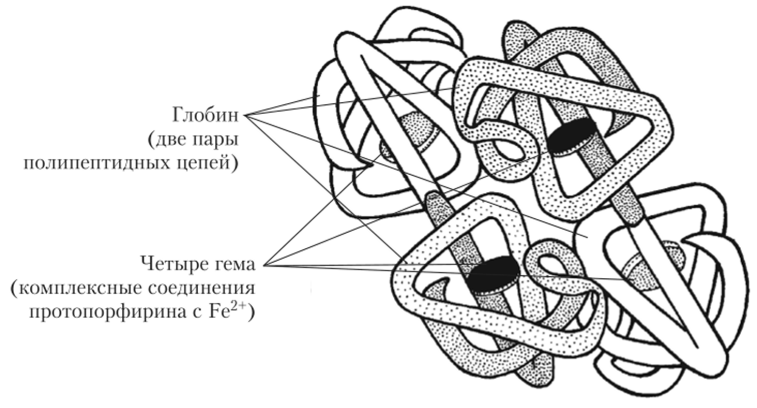
Рис. 4.4. Строение молекулы гемоглобина.
Гем представляет собой комплексное соединение протоиорфирина с двухвалентным железом. Ион Fe2+ обладает способностью обратимо связываться с кислородом. При этом железо не окисляется и остается двухвалентным. Гемоглобин, связанный с кислородом, называется оксигемоглобином. В артериальной крови содержание оксигемоглобина достигает 96—98% от общего количества гемоглобина (последнее составляет 130—170 г/л у мужчин и 120—150 г/л у женщин).
Вместе с тем, помимо кислорода, гемоглобин связывается с углекислым газом, образуя карбаминогемоглобин. Это свойство гемоглобина обеспечивает один из путей транспорта С02 кровью, а также снижает содержание угольной кислоты в плазме, участвуя в механизме поддержания pH крови на оптимальном уровне.
Соединения гемоглобина с кислородом и углекислым газом могут быть отнесены к «физиологическим» в противовес «патологическим» соединениям. Примером патологических соединений является карбоксигемоглобин, который образуется в результате взаимодействия гемоглобина с угарным газом (СО). Карбоксигемоглобин полностью лишен физиологических функций гемоглобина, что приводит к тяжелым отравлениям при поступлении в легкие угарного газа. Другим «патологическим» соединением гемоглобина является метгемоглобин, в котором железо окислено до трехвалентного состояния (Fe3+). Это лишает метгемоглобин способности обратимо связываться с кислородом. Небольшое количество метгемоглобина постоянно присутствует в крови. Перекисные соединения, свободные радикалы и другие окислители увеличивают долю метгемоглобина, антиоксиданты (например, аскорбиновая кислота) — снижают.
Тяжелые заболевания может вызвать не «дефект» гемоглобина, а его недостаточная концентрация. Патологические состояния, при которых в крови снижается количество гемоглобина, объединены под общим названием «анемия» (малокровие). Анемия может быть связана с острой кровоиотерсй. После такой кровонотери жидкая часть крови возмещается в течение одних-трех суток, а концентрация эритроцитов восстанавливается до нормы за три-шесть недель. Все это время организм страдает от анемии.
Причиной анемии может быть также нарушение функциональной активности красного костного мозга, связанное с его радиационным или химическим поражением. Такую анемию называют апластической. Репродукция эритроцитов в красном костном мозге может быть сильно замедлена при недостатке витамина В12 или фолиевой кислоты, в этом случае формируются слишком крупные красные клетки крови разнообразной формы (мегалобласты). Увеличение размеров не компенсирует замедления репродукции эритроцитов. Этот вид анемии называется мегалобластной анемией.
Анемию может вызвать синтез «дефектного»-гемоглобина, у которого повреждены р-цепочки. При низкой концентрации кислорода такой гемоглобин кристаллизуется. Кристаллы удлиняют эритроцит и придают ему вид серпа. Нетипичная форма приводит к быстрому разрушению эритроцитов при прохождении через капилляры и развитию анемии, которая получила название «серповидно-клеточная». Этим заболеванием страдают от 0,3 до 1,0% коренных жителей Западной Африки. Внешний вид эритроцитов при различных видах анемии показан на рис. 4.5.

Рис. 43. Изменение формы эритроцитов при разных типах анемии