Одномембранные органеллы.
Физиология растений в 2 т. Том 1

В результате появилась гипотеза об эндосимбиотическом происхождении этих органелл. Впервые она была высказана русским ботаником М. Д. Мережковским, а дальнейшее свое развитие получила в трудах немецкого ботаника А. Ф. В. Шимпера (1883). Согласно ей более миллиарда лет тому назад в гетеротрофные клетки примитивных эукариот в результате фагоцитоза смогли попасть свободно живущие цианобактерии… Читать ещё >
Одномембранные органеллы. Физиология растений в 2 т. Том 1 (реферат, курсовая, диплом, контрольная)
Эндоплазматическая сеть (эндоплазматический ретикулум, ЭПР; от лат. reticulum — сеть) (см. рис. 1.1) представляет собой сложную систему цистерн и каналов, отходящих от ядра и пронизывающих всю цитоплазму.
Цистерны — это расширения каналов. Стенки каналов состоят из мембран. Каналы и цистерны заполнены жидкостью — энхилемой, содержащей белки и другие соединения. На стенках некоторых каналов, обращенных в цитозоль, располагаются рибосомы, поверхность мембраны в этом месте становится шероховатой. Такие мембраны называют гранулярными. Поэтому выделяют два типа эндоплазматического ретикулума — шероховатый (или гранулярный) и гладкий, без рибосом. Присутствие рибосом на поверхности мембран временно. Мембраны шероховатого ЭПР, соединяясь с рибосомами, образуют полирибосомы на своей внешней стороне.
Эндоплазматический ретикулум участвует в синтезе белков, гликопротеидов и мембранных липидов. Шероховатый ЭПР — место синтеза мембранных белков, белков клеточной стенки или белков, переносимых в вакуоли. Белки цитозоля и ядра синтезируются на свободных полирибосомах (см. гл. 2). Синтезированные белки должны выделиться из клетки или попасть через мембрану ЭПР в его внутреннее пространство — энхилему. Попав во внутреннее пространство ЭПР, белки могут присоединять цепочки моносахаридов. Необходимые для синтеза мембранных липидов жирные кислоты поступают из пластид. В ЭПР жирные кислоты превращаются в ненасыщенные и встраиваются в мембраны. Кроме белков и липидов, в ЭПР синтезируется большинство вторичных метаболитов (сгеролы, флавоноиды, терпеноиды) (см. параграф 1.3). Вещества из ЭПР переносятся с помощью везикул к аппарату Гольджи.
ЭПР — один из клеточных депо, запасающих кальций, который может поступать в цитозоль в ответ на раздражение. Регулируя концентрацию кальция в цитозоле, ЭПР таким способом участвует в передаче раздражения в клетке (см. параграф 1.6). Разрушение вредных веществ, попавших в клетку, также одна из функций ЭПР.
Количество каналов и цистерн ЭПР зависит от физиологической активности клетки. В неблагоприятных условиях число цистерн увеличивается.
Аппарат Гольджи был обнаружен в клетках в 1898 г. итальянцем К. Гольджи значительно раньше рибосом. Каждый аппарат Гольджи состоит из 5—10 диктиосом (рис. 1.15), расстояние между которыми 20—25 нм.
Рис. 1.15. Аппарат Гольджи1.
Диктиосома — это плоский мешочек, тонкий в центре, толстый по периферии и окруженный мембраной. Между диктиосомами располагаются тонкие слои цитозоля. Кроме диктиосом, в состав аппарата Гольджи входит много везикул (от лат. visicula — пузырек) разного размера, которые отделяются от периферической части диктиосомы. В результате аппарат Гольджи (АГ) имеет два полюса: на одном в результате слияния везикул, транспортирующих вещества из ЭГ1Р, образуется новая диктиосома (проксимальный, или формирующийся, полюс), на противоположном полюсе диктиосома распадается на везикулы (дистальный, или зрелый, полюс). Оба процесса происходят одновременно. Таким образом, диктиосомы — образования временные.
Диктиосомы рассеяны по всему цитозолю либо по одиночке, либо группами. В одной клетке может находиться от 1 до 30 аппаратов Гольджи, а в железистых клетках — до нескольких сотен. При необходимости ЭПР может образовывать их заново.
Наиболее важная функция АГ — синтез олигои полисахаридов клеточной стенки и транспорт веществ в клетке с помощью везикул.
Постепенно образующиеся вещества транспортируются с помощью везикул от одной диктиосомы к другой и одновременно происходят их превращения. От последней диктиосомы везикулы отрываются и переносят образовавшиеся вещества к плазмалемме, клеточной стенке или к органеллам. Везикулы, покрытые специальным белком клатрином, направляются к вакуоли. Другие везикулы направляются к плазмалемме. Встраивание везикул в плазмалемму приводит к увеличению ее поверхности. Избыточный материал возвращается в цитозоль с помощью эндоцитоза (см. параграф 1.5). Находящиеся в везикуле молекулы целлюлозы выходят в нериплазматическое пространство и включаются в клеточную стенку. Так аппарат Гольджи участвует в образовании плазмалеммы и клеточной стенки.
Пероксисомы — это окруженные мембраной органеллы, диаметром 0,5—1,5 мкм, которые вместе с другими веществами содержат ферменты, участвующие в гликолатном цикле фотосинтеза (см. параграф 4.5).
В пероксисомах содержится фермент каталаза, разлагающий образующуюся перекись водорода, токсичную для клеток. В результате перекись водорода превращается в воду и одновременно окисляются вредные вещества: фенолы, муравьиная кислота, формальдегид, спирт, — поэтому эта реакция полезна для клетки. Токсичные органические вещества антропогенного происхождения, т. е. образующиеся в результате деятельности человека, назвали ксенобиотиками.
Пероксисомы находятся в клетках листьев, где образуют комплексы с хлоропластами и митохондриями.
Глиоксисомы — органеллы, окруженные одинарной мембраной и содержащие ферменты глиоксилатного цикла и ферменты, катализирующие (3-окислепие жирных кислот. С их помощью жирные кислоты превращаются в углеводы (см. параграфы 4.1 и 5.1).
Глиоксисомы возникают в клетках при прорастании семян, особенно масличных растений, и на свету с началом фотосинтеза у проростка превращаются в пероксисомы. Во время старения листа (при подготовке к листопаду) можно наблюдать обратное превращение пероксисом листа в глиоксисомы.
Пероксисомы и глиоксисомы возникают или в результате деления, или формируются заново из мембран эндоплазматического ретикулума. Последние исследования показали, что пероксисомы образуются из глиоксисом на свету в результате изменения ферментного состава.
Олеосомы — органеллы, содержащие растительные масла (от лат. oleum — масло) и находящиеся в цитозоле. В отличие от других органелл, олеосома покрыта мембраной, состоящей из мономолекулярного слоя полярных липидов, гидрофильные фосфатные группы которых обращены к цитозолю, а гидрофобные — внутрь олеосомы (см. параграф 1.4). В липидном монослое на поверхности олеосомы заякорены белки, которые называют олеозинами (рис. 1.16). Олеозины синтезируются в ЭПР. Они препятствуют слиянию олеосом в крупные образования во время прорастания семян при поступлении воды. Липидный монослой, покрывающий олеосому, содержит среди прочих белков липоксигеназы. Липоксигепазы — ферменты, участвующие в мобилизации запасных липидов олеосом при прорастании семян.[1]
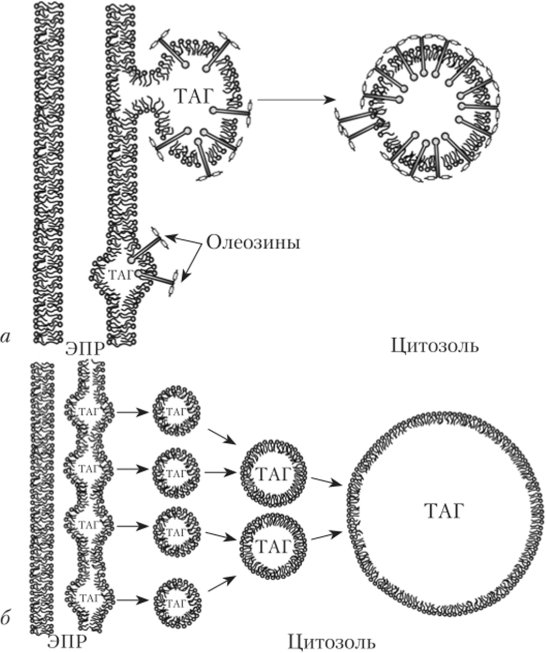
Рис. 1.16. Олеосомы растений1:
а — олеозин-зависимые олеосомы семян (0,2—1,0 мкм);
6 — олеозин-независимые олеосомы внесеменых частей (10—25 мкм); ЭПР — эндоплазматический ретикулум; ТАГ — триацилглицерины
Олеосомы формируются в эндоплазматическом ретикулуме за счет накопления молекул триацилглицеридов. Триацилглицериды — производные глицерина и жирных кислот, основной компонент мембран (см. параграф 1.4). Они синтезируются и накапливаются в мембранах ЭПР, толще липидного бислоя, до тех пор пока не будет достигнут окончательный размер олеосомы. Затем олеосома окружается фосфолипидным монослоем и отделяется от мембраны ЭПР.
Олеозины можно обнаружить только в олеосомах эндосперма, семядолей или зародыша.
Вакуоли (от лат. vacuum — пустой) характерны только для растительных клеток.
В меристематической клетке они маленькие и называются провакуолями. По мере роста клетки провакуоли сливаются друг с другом, и образуются несколько больших или одна центральная вакуоль (см. параграф 7.1). При этом из мембран мелких вакуолей формируется общая мембрана, отделяющая вакуоль от цитозоля и называемая тоиопласшом. В сформировавшейся клетке центральная вакуоль может занимать иногда более 90% ее объема.
Полость вакуоли заполнена клеточным соком, который содержит водорастворимые вещества: минеральные ионы (К4, С1~, Na+), аминокислоты, органические кислоты (малат, цитрат, оксалат) и их соли, сахара, некоторые низкомолекулярные белки, а также пигменты (антоцианы, беталаины), алкалоиды и другие вещества.
Вакуоль участвует в поступлении воды в клетку (см. параграф 3.2). За счет увеличения вакуоли происходит растяжение и увеличение размера самой клетки. Следовательно, благодаря вакуоли сильно увеличивается поверхность клетки. Такой способ увеличения поверхности более выгоден, чем, например, увеличение объема цитозоля, так как в последнем случае был бы необходим энергетически затратный синтез метаболитов, прежде всего белков, а также дополнительное количество азота — самого дефицитного элемента. Увеличение же вакуоли связано с поступлением воды, которое требует значительно меньшего расхода энергии.
В вакуолях откладываются запасные вещества, которые при необходимости могут возвращаться обратно в цитозоль, хранится запас нитратов и фосфатов, углеводы, в вакуолях листа может накапливаться малат (см. параграф 4.5). В результате вакуоль помогает поддерживать постоянное количество сахаров и ионов в цитозоле.
В вакуолях созревающих зерновок злаков и в семядолях бобовых растений откладываются запасные белки, в результате образуются алейроновые зерна (от греч. aleuron — пшеничная мука).
В вакуоли поступают и конечные продукты обмена веществ, которые не играют никакой роли в метаболизме и были бы вредны для цитоплазмы и ее ферментов. Некоторые из этих веществ — яды. Поступая в вакуоль, они исключаются из метаболизма. В вакуоль выделяются и другие ненужные для клетки вещества, например тяжелые металлы, здесь они перерабатываются и обезвреживаются. Поэтому еще одна важная функция вакуоли — изоляция и обезвреживание токсичных веществ.
Растворенные в клеточном соке пигменты, окрашивающие цветки и плоды, служат для привлечения насекомых-опылителей, а также птиц и животных — распространителей плодов и семян. Некоторые водорастворимые пигменты поглощают УФ-лучи и таким образом защищают фотосинтетический аппарат и мембраны от разрушения. Вместе с тем, содержащиеся в вакуоли фенольные соединения, алкалоиды, изоиреноиды, цианогеиные гликозиды, а также ферменты (хитиназы, глюкаиазы), разрушающие клеточные стенки грибов, насекомых, защищают растения от патогенов и травоядных животных.
Вакуоли возникают из цистерн эндоплазматического ретикулума.
Одним из механизмов образования вакуолей является автофагия (от греч. autos — сам и phagos — пожирающий) — особый механизм формирования вакуолей. Часть канала гладкого ЭПР образует пальцеобразные выросты, которые как щупальцы захватывают кусочек цитоплазмы. Когда соседние щупальца смыкаются, образуется автофагосома, окруженная сначала двойной мембраной. Затем внутренняя мембрана разрушается, а наружная превращается в тонопласт. Автофагосома содержит много кислых гидролитических ферментов. В нее поступает вода, и вакуоль увеличивается.
Эндоплазматический ретикулум, аппарат Гольджи и вакуоль вместе образуют эндомембраниую систему клетки.
Лизосомы представляют собой маленькие везикулы (диаметром около 1 мкм), окруженные мембраной. Форма и размеры лизосом различны. Внутри находится энхилема, содержащая гидролитические ферменты. Образующиеся в результате действия этих ферментов аминокислоты и углеводы транспортируются в цитозоль и могут вторично использоваться. В определенные моменты жизни клетки эти ферменты могут выходить из лизосомы и разрушать внутриклеточные структуры, выполнившие свои функции.
Образование и функционирование лизосом связано с эндоплазматической сетью и аппаратом Гольджи.
В последнее время некоторые авторы стали считать, что лизосомы есть только в клетках грибов и животных, а в растительных клетках их роль выполняют специальные вакуоли, названные литическими. Литические вакуоли отличаются от запасающих вакуолей кислотностью клеточного сока: запасающие вакуоли имеют нейтральный pH, а литические — кислый pH.
Итак, каждая органелла занимает в клетке определенное положение. В разное время своей жизни клетка имеет различное строение. (Сравните строение, например, паренхимной клетки с меристематической.) Поэтому мы можем сказать, что клетка организована в пространстве и во времени.
Протопласт в целом и все органеллы построены по общему принципу. Протопласт окружен нлазмалеммой; ядро, пластиды, митохондрии окружены двойной мембраной; аппарат Гольджи, эндоплазматический ретикулум, вакуоли, пероксисомы, лизосомы — одинарной. Только рибосомы, микротрубочки и микрофиламенты не имеют мембран. Наличие в клетке большого количества мембран позволяет заключить, что как организм состоит из клеток, так клетка состоит из мембран, или по-другому: мембрана является универсальной структурной единицей клетки. Каждая органелла и клетка в целом имеют свою внутреннюю среду — строму, матрикс или энхилему.
Итак, клетка представляет собой систему — сложный комплекс элементов (органелл), расположенных в определенном порядке, связанных друг с другом и выполняющих определенные функции в соответствии с программой этой системы. Впервые предложил рассматривать клетку как систему в 1932 г. Л. фон Берталанфи (Канада).
Пластиды и митохондрии занимают особое положение в клетке среди других органелл: они окружены двумя мембранами, отличающимися по проницаемости для веществ, а также имеют собственную систему синтеза белка, способ деления у них такой же, как и у бактерий. Липиды и белки внутренней мембраны митохондрий по составу похожи на липиды и белки бактерий. Синтез белка в митохондриях и бактериях ингибируется одними и теми же антибиотиками, которые не влияют на синтез белка в других частях растительной клетки. Они имеют геном, похожий на геном цианобактерий, в митохондриях содержатся плазмиды — микрокольцевые молекулы ДНК, способные к саморепликации.
В результате появилась гипотеза об эндосимбиотическом происхождении этих органелл. Впервые она была высказана русским ботаником М. Д. Мережковским, а дальнейшее свое развитие получила в трудах немецкого ботаника А. Ф. В. Шимпера (1883). Согласно ей более миллиарда лет тому назад в гетеротрофные клетки примитивных эукариот в результате фагоцитоза смогли попасть свободно живущие цианобактерии в качестве внутренних симбионтов, которые затем постепенно превратились в митохондрии и хлоропласты. Наличие в клетке трех относительно самостоятельных геномов — ядерного (хромосомного), иластидного и митохондриального — является следствием эндосимбиотического происхождения растительной клетки.
Клетка обладает всеми свойствами живой материи — обменом веществ, ростом, размножением, движением, раздражимостью, способностью приспосабливаться к изменяющимся окружающим условиям.
Контрольные вопросы
- 1. Каково строение клеточной стенки и из каких основных веществ она состоит? Каковы ее свойства и функции?
- 2. Что называют свободным пространством клетки? Каковы его функции?
- 3. Что такое плазмодесмы? Каковы их строение и функции?
- 4. Что такое симпласт, апопласт?
- 5. Что такое протопласт? В чем состоят особенности его химического состава?
- 6. Каковы строение и функции ядра?
- 7. Какие органеллы входят в состав цитоплазмы?
- 8. Что такое цитозоль? Каков его химический состав, свойства и функции?
- 9. Из чего состоит цитоскслет?
- 10. Что такое микрофиламенты?
- 11. Какие функции выполняет аппарат Гольджи?
- 12. Какие процессы идут в эндоплазматическом ретикулуме?
- 13. Какую роль играет вакуоль в клетке?
- 14. Какие органеллы не окружены мембраной?
- 15. Какие функции выполняют микротрубочки? Из чего они состоят?
- 16. Каковы строение и функции митохондрии? Как она образуется?
- 17. Какое строение имеет гранальный хлоропласт?
- 18. Какое строение у агранального хлоропласта?
- 19. Как образуются хлоропласта? Какими свойствами и функциями они обладают?
- 20. Каковы строение и функции пероксисомы?
- 21. Что такое лизосома? Какие ферменты она содержит?
- 22. Что такое олеосомы?
- 23. Что является внутренней средой клетки, органеллы?
- 24. Какими свойствами живой материи обладает клетка?
- 25. Какую роль играет движение цитозоля и органелл?
- 26. Что вы можете рассказать о движении цитоплазмы?
- [1] Цит. по: Chapman К. D., DyerJ. М., Mullen R. Т. Biogenesis and functions of lipid dropletsin plants//J. Lipid Research. 2012. V. 53. P. 215−226.