Растительные ткани (гистология)

Членики ситовидных трубок формируются из вытянутых клеток прокамбия или камбия. При этом материнская клетка меристемы делится в продольном направлении и производит две клетки. Одна из них превращается в членик, другая — в клетку-спутницу. Наблюдается и поперечное деление клетки-спутницы с последующим образованием двух-трех подобных клеток, расположенных продольно одна над другой рядом с члеником… Читать ещё >
Растительные ткани (гистология) (реферат, курсовая, диплом, контрольная)
У большинства наземных растений клетки тела неодинаковы по своим функциям, строению и отчасти происхождению. Эта разнородность возникла и закрепилась в процессе приспособления растений к различиям воздушной и почвенной сред.
Системы клеток, структурно и функционально сходные друг с другом и обычно имеющие общее происхождение, получили название тканей.
Ткани есть почти у всех растений. Нет их только у части моховидных (большинство печеночников). В отличие от животных растения растут и образуют новые клетки, ткани, органы на протяжении всей жизни, то есть относятся к организмам с открытым ростом. Индивидуальное развитие растений начинается со стадии зародыша. Большинство клеток зародыша обладает, так называемой, меристематической активностью, то есть способностью к делению. Часть из них сохраняет эту способность в течение всей жизни. Они получили название инициальных клеток, или инициален Другая часть клеток, постоянно возникающих в процессе деления инициален, получила название производных. Эти клетки обычно делятся один или два раза и начинают дифференцироваться в специализированные, обычно лишенные способности к делению, типы клеток. Сам процесс дифференцировки — это превращение инициалей и их производных в специализированные клетки, выполняющие строго определенные функции.
Существуют различные классификации тканей. Все они достаточно условны и используются по преимуществу для удобства обзора и с целью облегчения обучения. Чаще растительные ткани делят на несколько групп в зависимости от основной функции: 1) меристемы, или образовательные ткани; 2) покровные; 3) механические; 4) проводящие; 5) основные; 6) секреторные, или выделительные.
Ткани, состоящие их одного типа клеток, получили название простых, а состоящие из разных типов — сложных, или комплексных.
Образовательные ткани: особенности строения, типы меристем, их функциональные свойства и локализация в органах растений. Меристемы, или образовательные ткани, обладают способностью к делению и образованию новых клеток, занимая ничтожный объем в теле растения (примерно 0,1% общей массы). Меристемы формируют все прочие ткани и определяют длительный (в течение всей жизни) рост растения. Инициальные клетки меристем задерживаются на эмбриональной стадии развития в течение всей жизни растения, а их производные постепенно дифференцируются и превращаются в клетки различных постоянных тканей. Тело наземных растений — производное относительно немногих инициальных клеток.
По локализации в теле растения различают верхушечные (апикальные), боковые (латеральные), вставочные (интеркалярные) и раневые (травматические) меристемы (табл. 4).
Типы меристем по локализации в теле растения и их характеристика.
Апикальные (верхушечные) | Латеральные (боковые) | Интеркалярные (вставочные) | Раневые |
Располагаются на верхушках корней и побегов, формируя конусы нарастания корней и побегов и обеспечивая их нарастание в длину. По происхождению первичны. | Располагаются параллельно боковым поверхностям осевых органов, нередко образуя цилиндры, на поперечных срезах имеющие вид колец. Боковые меристемы обеспечивают утолщение корня и стебля. По происхождению могут быть:
Из прокамбия и камбия образуются проводящие ткани, из феллогена — перидерма. | Сохраняются в виде отдельных участков в зонах активного роста, например, у оснований междоузлий и в основании листьев злаков. По происхождению первичны. | Образуются из клеток основной паренхимы в местах повреждения тканей и органов, дают начало каллусу — особой ткани, состоящей из однородных тонкостенных паренхимных клеток, прикрывающих место поражения. Таким образом, эта меристема является вторичной по происхождению. При пропитывании клеточных оболочек каллуса суберином формируется пробка. Каллус и пробка защищают травмированный участок от поражения патогенами. |
Цитологические особенности меристем. Наиболее выражены у апикальных меристем. Клетки — изодиаметрические многогранники — не разделены межклетниками. Клеточные стенки тонкие, с малым содержанием целлюлозы. Цитоплазма густая, ядро крупное, расположено в центре. Вакуоли многочисленные, мелкие. Эргастические вещества, как правило, отсутствуют. Пластид и митохондрий мало, и они мелкие (рис. 95).
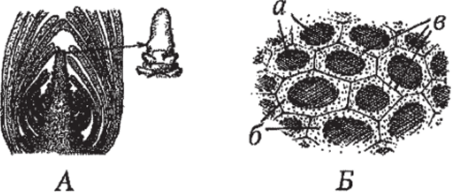
Рис. 95. Строение конуса нарастания и клеток апикальной меристемы:
А — конус нарастания апикальная меристема): продольный разрез и внешний вид; Б — клетки апикальной меристемы: а — ядра; б — густая цитоплазма; в — тонкие клеточные стенки и срединные пластинки.
Покровные ткани. Располагаются на границе с внешней средой, то есть на поверхности органов растения. Большинство из них состоит из плотно сомкнутых живых, реже мертвых клеток. Они выполняют барьерную роль, защищая внутренние ткани от высыхания и повреждения. Одна из функций покровных тканей побега — регуляция газообмена и транспирации. Некоторые из них способны к всасыванию и выделению, активно регулируя скорость и избирательность проникновения веществ. Кроме того, покровные ткани — достаточно серьезный барьер для проникновения патогенных микроорганизмов.
Принято различать первичные покровные ткани, образующиеся в результате дифференцировки клеток первичных меристем, эпидерму й эпиблему. Вторичная покровная ткань — перидерма — образуется из вторичной меристемы — феллогена.
Первичные покровные ткани: характеристика эпидермы и эпиблемы. Сравнительная характеристика первичных покровных тканей представлена в таблице 5.
Сравнительная характеристика эпидермы и эпиблемы.
Jfen/n. | Признак. | Эпидерма (эпидермис, кожица). | Эпиблема (ризодерма). |
1. | На поверхности какого органа располагается? | Первичная покровная ткань молодых зеленых побегов, а также листьев и плодов. | Первичная однослойная покровная ткань корня. |
2. | Из чего образуется? | Из первичной меристемы — протодермы конуса нарастания побега. | Из протодермы — наружного слоя клеток апикальной меристемы корня. |
3. | Особенности клеток. | Клетки плотно сомкнуты, нередко с извилистыми оболочками, благодаря чему повышается плотность их сцепления друг с другом. Это живые клетки с тонким постенным слоем протопласта с мелкими редкими лейкопластами и ядром. Имеют одну крупную вакуоль с бесцветным клеточным соком. | Клетки тонкостенные, лишенные кутикулы, с вязкой цитоплазмой, с большим числом митохондрий (активное поглощение веществ происходит с затратой энергии). Каждая клетка эпиблемы способна к образованию корневого волоска. |
4. | Толщина клеточных стенок. | Характерная особенность клеток эпидермы — неодинаковая толщина стенок. Наружные стенки более толстые, часто покрыты кутикулой или воском. Обычно кутикула лежит тонким ровным слоем, иногда она местами образует утолщения в виде складок (складчатая кутикула). | Стенки тонкие. |
5. | Наличие устьиц. | ; | |
6. | Функции. | Регуляция газообмена и транспирации, то есть испарения воды растением. | Осуществление поглощения воды и минеральных солей из почвы. |
Устьица представляют собой высокоспециализированные образования эпидермы через которые осуществляется транспирация и газообмен. Состоят устьица из двух замыкающих клеток и устьичной щели между ними (рис. 96). Щель может расширяться и сужаться и, тем самым, регулировать транспирацию и газообмен. Под ней располагается воздушная полость, окружённая мякотью листа. Клетки эпидермы, примыкающие к замыкающим, называются побочными (сопровождающими, околоустьичными). Они участвуют в движении замыкающих клеток и вместе с ними образуют устьичный аппарат. Механизм движения замыкающих клеток весьма сложен и неодинаков у разных видов. У большинства растений при недостаточном водоснабжении в ночные часы, а иногда и днём, тургор в замыкающих клетках понижается и щель замыкается, снижая тем самым уровень транспирации. С повышением тургора устьица открываются. Считают, что главную роль в этих изменениях принадлежат ионам калия. Существенное значение в регуляции тургора имеет присутствие в замыкающих клетках хлоропластов. Первичный крахмал хлоропластов, превращаясь в сахар, повышает концентрацию клеточного сока. Это способствует притоку воды из соседних клеток и переходу замыкающих клеток в упругое состояние, вследствие чего устьице открывается. В темноте при недостаточном обводнении устьичная щель закрывается из-за понижения тургора в замыкающих клетках. Устьица обычно располагаются на нижней стороне листа. Однако у водных растений, обладающих плавающими листьями, они находятся только на верхней стороне листа.
По числу сопровождающих клеток и их расположению относительно устьичной щели устьица подразделяются на ряд типов (рис. 97). У аномоцитного типа устьичного аппарата побочные клетки не отличаются от остальных клеток эпидермы. Диацитный тип характеризуется только двумя побочными клетками, общая стенка которых перпендикулярна устьичной щели (сем. Губоцветные). При парацитном типе побочные клетки располагаются параллельно замыкающим и устьичной щели (хвощи). Анизоцитный тип имеет три побочные клетки, одна из которых заметно крупнее или мелме остальных (сем. Крестоцветные). Тетрацитный тип отличается четырьмя побочными клетками (характерен для однодольных растений). При энциклоцитном типе побочные клетки образуют узкое кольцо вокруг замыкающих клеток.
Наряду с устьицами, предназначенными для газообмена и транспирации, существуют особые устьица, завершающие гидатоды и предназначенные для выделения капельно-жидкой воды.
У некоторых растений под эпидермой расположена особая ткань — гиподерма. Она отчасти выполняет механическую функцию — отчасти предохраняет растение от избыточного испарения. Хорошо развитая гиподерма заметна в своеобразных игольчатых листьях — хвоинках сосны. Располагаясь непосредственно под эпидермой, гиподерма формируется независимо от неё.
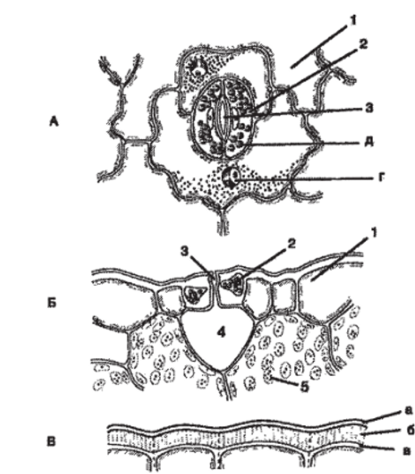
Рис. 96. Строение устьица: А, Б — кожица листа тимьяна (вид сверху и на поперечном срезе); В — кожица со стебля цереус (сем. кактусовых);
- 1 — собственно эпидермальные клетки; 2 — замыкающие клетки устьица;
- 3 — устьичная щель; 4 — воздухоносная полость;
- 5 — клетки хлорофиллоносной паренхимы; а — кутикула; б — кутикулярный слой — оболочка с суберином и воском; в — целлюлозный слой стенки; г — ядро с ядрышком; д — хлоропласты
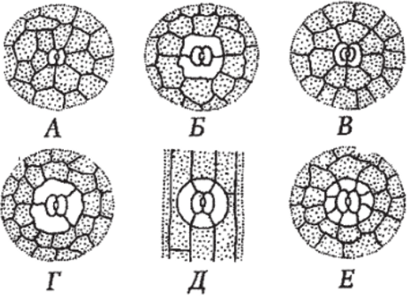
Рис. 97. Основные типы устьичных аппаратов: А — аномоцитный; Б-диацитный; В — парацитнмй; Г- анизоцитный; Д-тетрацитный; Е — энциклоцигный.
Трихомы — это различные по форме, строению и функциям выросты клеток эпидермы: волоски, чешуйки, щетинки и т. п. Они подразделяются на кроющие и железистые. Железистые трихомы в отличие от кроющих имеют секреторные клетки с секретом, являющимся продуктом выделения (см. Секреторные ткани). Кроющие волоски образуют на растении шерстистый, войлочный или иной покров, отражая часть солнечных лучей, чем уменьшают транспирацию. То есть совокупность волосков образует опушение, которое, в зависимости от густоты, может быть рассеянным, плотным, войлочным, шерстистым и др. Иногда волоски располагаются только там, где расположены устьица, например на нижней стороне листа мать-и-мачехи. У некоторых растений живые волоски увеличивают общую испаряющую поверхность, что способствует ускорению транспирации. Размеры трихом варьируют в значительных пределах. Наиболее длинные трихомы (до 5−6 см) покрывают семена хлопчатника. Кроющие трихомы (волоски) могут быть однои многоклеточными, звездчатыми, ветвистыми, Т-образными и др., с тонкими или толстыми стенками, с гладкой или бородавчатой поверхностью (рис. 98). Кроющие трихомы могут длительное время оставаться живыми или быстро отмирать и заполняться воздухом.
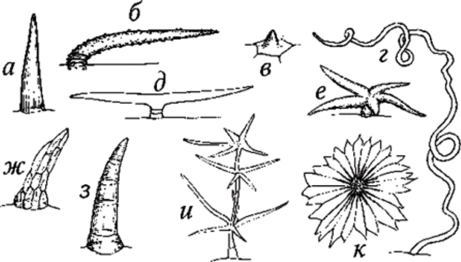
Рис. 98. Простые волоски: а — одноклеточный с гладкой кутикулой; б — двух-трехклеточный прижатый с бородавчатой кутикулой; в — сосочковидный; г - одноклеточный бичевидный; д — двух-трехклеточный Т-образный; е — многоклеточный звездчатый; ж — пучковый; з — многоклеточный с гладкой кутикулой; и — многоклеточный ветвистый; к — многоклеточный пельтатный (щитовидный) От трихом, возникающих только при участии эпидермальных клеток, отличаются эмергенцы, в образовании которых участвуют и более глубоко расположенные ткани субэпидермального слоя (рис. 99).
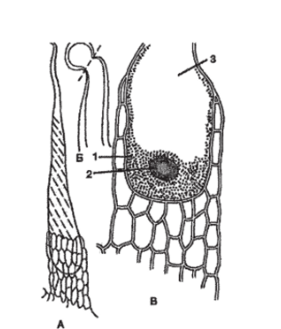
Рис. 99. Эмергенцы (у крапивы): А — общий вид;
Б — окончания волоска (показана линия облома);
В — основание волоска с цитоплазмой (1), ядром (2) и вакуолью (3).
Эпидерма двудольных и однодольных имеет различное строение: клетки эпидермы двудольных растений паренхимные, устьица расположены в разные стороны; клетки эпидермы однодольных растений имеют прозенхимную форму, устьица расположены параллельно друг другу (рис. 100).
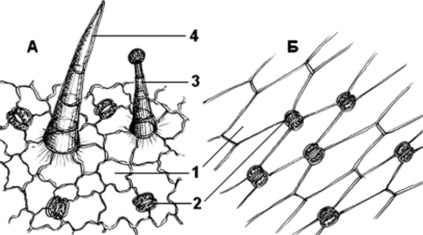
Рис. 100. Эпидерма: А двудольных; Б — однодольных: 1 — клетки эпидермы;
2 — устьица; 3 — головчатый (железистый) волосок; 4 — простой волосок.
Вторичные покровные ткани, образование, строение, функции. Перидерма — это сложная многослойная вторичная покровная ткань стеблей, корней и корневищ двудольных многолетних (реже однолетних) растений. Она сменяет постепенно отмирающие первичные покровные ткани осевых органов. Обычно перидерма появляется в течение первого вегетационного периода в тех частях корня и побега, которые прекратили рост в длину. Ев образование обусловлено деятельностью вторичной меристемы — феллогена (пробковый камбий). Эта деятельность сводится к делению клеток в центробежном направлении и дифференциации их в пробку (феллему), а также в центростремительном направлении и превращении в живые паренхимные клетки феллодермы. Феллоген может закладываться в эпидерме стеблей в субэпидермальном слое. Таким образом, перидерма — это комплекс, состоящий из феллемы (покровная ткань, пробка), феллогена (образовательная ткань) и феллодермы (основная ткань, хлорофиллоносная паренхима).
Пробка состоит из отмерших клеток, лишённых межклетников (рис. 101). Их оболочки пропитаны суберином. Клетки пробки воздухеи водонепроницаемы. Они защищают живые ткани от чрезмерного испарения, резких колебаний температуры, проникновения болезнетворных микробов. Живые ткани, лежащие под пробкой, нуждаются в газообмене и удалении избытка влаги. Это достигается с помощью образующихся в перидерме чечевичек — участков с рыхло располагающимися клетками (выполняющая ткань чечевички) (рис. 101). К зиме они закрываются тонким слоем замыкающих клеток, который весной при возобновлении деятельности феллогена разрывается. По мере утолщения ветвей растения чечевички растягиваются. Чечевички, имеющие вид небольших бугорков, отчетливо видны на поверхности побегов деревьев и кустарников. На стволах березы их остатки наблюдаются в виде характерных поперечных черных полосок, у осины они принимают форму ромбов.
Корка, образование, строение, функции В многолетних осевых органах растений обычно развивается несколько перидерм. Причем каждая последующая закладывается глубже предыдущей. Со временем клетки наружных перидерм и располагающиеся между ними ткани отмирают, в результате чего образуются мощный покровный комплекс — корка Корка ежегодно наращивается за счет заложения новых слоев перидермы (рис. 101).
Корка не способна к растяжению, поэтому при утолщении ствола в ней появляются трещины. На дне трещин во внутренней перидерме имеются чечевички, обеспечивающие газообмен. Корка надежно предохраняет стволы от механических повреждений, лесных пожаров, резкой смены температуры.
Механические ткани, их виды, особенности строения и локализация в органах растений. Механические ткани — это опорные (арматурные) ткани, образующие скелет растения и обеспечивающие его прочность, вследствие чего растение способно противостоять нагрузкам на растяжение, сжатие и изгиб. Механические ткани наиболее развиты в осевой части побега — стебле. Они чаще располагаются по его периферии отдельными участками или сплошным кольцом, чем достигается наибольший защитный эффект при изгибе органа. В корне же, напротив, механические ткани располагаются в центре, что позволяет выдерживать сопротивление на разрыв. Механические ткани образуются как из первичных, так и из вторичных меристем. Наиболее заметная особенность клеток механических тканей — их значительно утолщенные оболочки, которые продолжают выполнять опорную функцию даже после отмирания их живого содержимого. Различают механические ткани с равномерно и неравномерно утолщенными клеточными стенками. Выделяют три основных типа механических тканей: колленхиму, склеренхиму и склереиды.
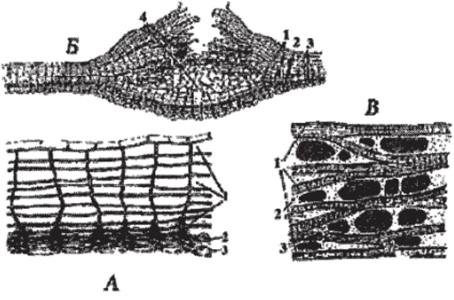
Рис. 101. Вторичные покровные ткани и их элементы:
А — строение перидермы: 1 — пробка (сверху остатки эпидермы); 2 — феллоген; 3 — феллодерма; Б — строение чечевички: 1 — пробка (сверху остатки эпидермы); 2 — феллоген; 3 — феллодерма; 4 — выполняющая ткань; В — схема строения корки: 1 — слои перидермы; 2 — отмершая основная ткань;
3 — группы механических элементов (волокон и (или) склереид) Виды механических тканей и их характеристика Таблица 6.
Признак. | Вид механической ткани. | ||
Колленхима. | Склеренхима. | Склереиды. | |
Происхождение. | Первичное. | Первичное и вторичное. | Первичное. |
Форма клеток. | Прозенхимная (клетки вытянуты вдоль оси органа). | Прозенхимная. | Паренхимная. |
Состояние клеток. | Живые, выполняют опорную функцию в состоянии тургора. | Мертвые, на определенном этапе дифференциации лишаются протопласта. | Мертвые. |
Клеточные оболочки. | Первичные неравномерно утолщенные неодревесневающие. | Вторичные равномерно утолщенные одревесневающие, реже неодревесневающие. | Вторичные равномерно утолщенные одревесневающие. |
В каких органах встречаются. | Молодые стебли и черешки листьев. | Стебли, корни, листья. | В виде скоплений или поодиночке (клетки идиобласты) в плодах, семенах, листьях, стеблях. |
В зависимости от характера утолщений стенок и соединений клеток между собой различают три типа колленхимы: уголковую, пластинчатую и рыхлую. Уголковая колленхима имеет стенки, утолщенные в углах клеток. Утолщения стенок соседних клеток смыкаются, образуя трех-, четырехили пятиугольники (рис. 102). Часто располагается под эпидермой над главной жилкой листьев, а также в форме продольных тяжей в выступающих ребрах стеблей травянистых растений. Пластинчатая колленхима имеет утолщенные тангентальньге стенки клеток. Радиальные стенки у нее остаются тонкими. Часто пластинчатая колленхима образует в стебле сплошное кольцо, располагаясь непосредственно под эпидермой. Рыхлая колленхима в отличие от первых двух имеет хорошо выраженные межклетники. Утолщению подвергаются лишь те части оболочек, которые прилегают к межклетным пространствам.
Клетки колленхимы, будучи живыми с неодревесневшими стенками, способны к росту в длину и не препятствуют росту органов, в которых они располагаются. Иногда колленхима содержит хлоропласты.
Склеренхимные клетки, часто называемые волокнами, подразделяют на: лубяные — волокна, входящие в состав флоэмы (луба) и древесинные (волокна либриформа) — входящие в состав ксилемы (рис. 103). Лубяные волокна сильно вытянутые прозенхимные клетки с заостренными концами, в исключительных случаях достигают несколько десятков сантиметров длины (например, у рами). Их равномерно утолщенные вторичные клеточные оболочки очень часто не одревесневают, а остаются целлюлозными. Неодревесневающие волокна стеблей двудольных используют для изготовления различных тканей (волокна льна, рами, кенафа), реже веревок (пенька, получаемая из конопли). Помимо луба могут встречаться в листовых черешках и пластинках, в цветоножках, реже в плодах. Древесинные волокна (волокна либриформа) короче лубяных (не более 2 мм) и их стенки всегда одревесневают. Они имеют скошенные концы, оболочки таких волокон обладают прочностью, близкой к прочности стали. Поры в оболочках немногочисленные, простые.
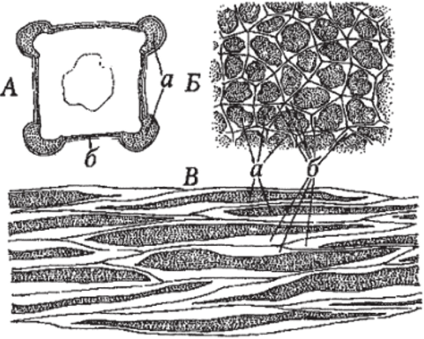
Рис. 102. Колленхима:
А — локализация видов колленхимы в стеблях губоцветных: а — уголковая; б — пластинчатая; Б — вид уголковой колленхимы на поперечном срезе, В — вид уголковой колленхимы на продольном срезе: а — протопласты, б — первичная клеточная стенка, утолщенная в углах клеток.
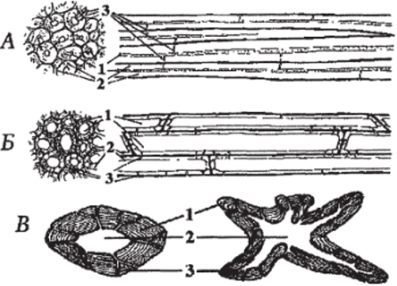
Рис. 103. Склеренхима и склереиды:
А — лубяные волокна в продольном (справа) и поперечном (слева) разрезах, Б — древесинные волокна в продольном (справа) и поперечном (слева) разрезах:
1 — полость; 2 — толстая вторичная клеточная стенка; 3 — простые поры;
В — склереида в разрезе, Г — ветвистая склереида в разрезе:
- 1 — толстая одревесневшая вторичная клеточная стенка;
- 2 — полость или протопласт; 3 — ветвящиеся простые поры
Склереиды — структурные элементы механической ткани, обычно возникают из клеток основной паренхимы в результате утолщения и лигнификации их оболочек. Зрелые склереиды сильно варьируют по форме (рис. 103). Склереиды могут встречаться в виде скоплений, либо располагаться поодиночке (клетки идиобласты). Считается, что функция склереид — противостоять сдавливанию, но иногда они защищают зеленые части растений от поедания.
Проводящие ткани: особенности строения, классификация, функции. Проводящие ткани служат для передвижения (транспорта) по растению растворенных в воде питательных — органических и неорганических веществ. Подобно покровным тканям, они возникли как следствие приспособления растений к жизни в двух средах: почвенной и воздушной. В связи с этим появилась необходимость транспортировки питательных веществ в двух направлениях. От корня к листьям движется восходящий, или транспирационный, ток водных растворов солей, который осуществляется почти исключительно по трахеальным элементам ксилемы. Ассимиляционный, нисходящий, ток органических веществ направляется от листьев к корням и осуществляется по ситовидным элементам флоэмы. Проводящие ткани объединяют все органы растения в единую систему. Помимо дальнего, т. е. осевого, транспорта питательных веществ, по проводящим тканям частично осуществляется и средний — радиальный транспорт.
Все проводящие ткани являются сложными, или комплексными, т. е. состоят из морфологически и функционально разнородных элементов (табл. 7).
Таблица 7.
Элементы ксилемы и флоэмы.
Проводящая ткань. | Проводящие элементы. | Механические элементы. | Основные элементы. |
Ксилема. | Трахеиды (есть у спорофитов всех растений, а у большинства хвощевидных, плауновидных, папоротниковидных и голосеменных они являются единственным проводящим элементом ксилемы) и сосуды | Древесинные волокна (волокна либриформа). | Древесинная паренхима (служит местом хранения запасных веществ, также по ней осуществляется средний транспорт). |
Флоэма. | Ситовидгше клетки (основной проводящий элемент флоэмы у всех групп растений, исключая покрытосеменные) юм ситовидные трубки с клетками-спутницами (проводящий элемент флоэмы покрытосеменных). | Лубяные волокна | Лубяная паренхима (служит местом хранения запасных веществ, также по ней осуществляется средний транспорт). |
Ксилема: ее основные элементы, их строение и функции. Термин ксилема ввел немецкий ботаник К. В. Негели в 1858 г. Первичная и вторичная ксилемы содержат клетки одних и тех же типов. Однако первичная ксилема не имеет сердцевинных лучей, отличаясь этим от вторичной. Первичная ксилема формируется из прокамбия, вторичная — из камбия. В состав ксилемы входят морфологически и функционально различные элементы, осуществляющие функции проведения, хранения веществ, а также чисто опорные функции (табл. 6). Дальний транспорт осуществляется по трахеальным элементам ксилемы: трахеидам и сосудам.
Трахеиды в зрелом состоянии — это мертвые прозенхимные клетки, суженные на концах и лишенные протопласта (рис. 104). Длина трахеид в среднем составляет 1−4 мм, поперечник же не превышает десятых и даже сотых долей миллиметра. Стенки трахеид одревесневают, утолщаются и несут простые или окаймленные поры, через которые происходит фильтрация растворов, с помощью которой осуществляется дальний транспорт. Большая часть окаймленных пор находится около окончаний клеток, то есть там, где растворы «просачиваются» из одной трахеиды в другую.
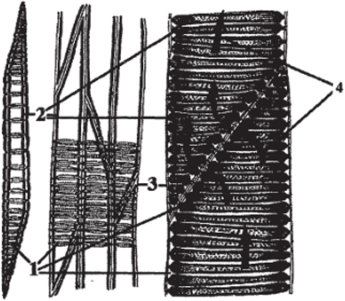
Рис. 104. Трахеиды: Л — общий вид трахеиды с окаймленными порами;
Б — вид стыка двух трахеид в разрезе на большом увеличении:
- 1 — окаймленные поры; 2 — толстая вторичная одревесневшая клеточная стенка в виде обручей; 3 — тонкая первичная неодревесневшая клеточная стенка;
- 4 — движение воды
Сосуды — это полые трубки, состоящие из отдельных члеников, располагающихся друг над другом. Между расположенными одним над другим члениками одного и того же сосуда имеются разного типа сквозные отверстия — перфорации. Благодаря перфорациям вдоль всего сосуда свободно осуществляется восходящий ток жидкости. Эволюционно сосуды, по-видимому, произошли из трахеид путем разрушения замыкающих плёнок пор и последующего их слияния в одну или несколько перфораций. Концы трахеид, первоначально сильно скошенные, заняли горизонтальное положение, а сами трахеиды стали короче и превратились в членики сосудов. Сосуды могут состоять из весьма значительного числа члеников, различной длины и диаметра. Общая же длина сосудов достигает иногда нескольких метров. Помимо первичной оболочки, сосуды, как и многие трахеиды, в большинстве случаев имеют вторичные утолщения. В самых ранних трахеальных элементах вторичная оболочка может иметь форму колец, не связанных друг с другом (кольчатые сосуды и трахеиды). Позднее появляются трахеальные элементы со спиральными утолщениями. Затем следуют сосуды и трахеиды с утолщениями, которые могут быть охарактеризованы как спирали, витки которых связаны между собой (лестничные утолщения). В конечном итоге вторичная оболочка сливается в более или менее сплошной цилиндр, формирующийся внутрь от первичной оболочки. Этот цилиндр прерывается в отельных участках порами. Сосуды и трахеиды с относительно небольшими округлыми участками первичной клеточной оболочки, не прикрытыми изнутри вторичной оболочкой, нередко называют пористыми (рис. 105). Вторичная, а иногда и первичная оболочки, как правило, лигнифицируются, то есть пропитываются лигнином, что придаёт им дополнительную прочность, но ограничивает возможности дальнейшего их роста в длину.

Рис. 105. Сосуды: 1 — кольчатый; 2 — спиральный; 3 — лестничный;
4 — сетчатый; 5 — точечный (пористый).
Флоэма: ее основные элементы, их строение и функции. Термин «флоэма» ввел К. В. Негели в 1858 г. Флоэма — сложная проводящая ткань, по которой осуществляется транспорт продуктов фотосинтеза от листьев к местам их использования или отложения (к точкам роста, подземным органам, зреющим семенам и плодам и т. д.).
Первичная флоэма дифференцируется из прокамбия, вторичная (луб) — производное камбия. В стеблях флоэма располагается обычно снаружи от ксилемы, а в листьях обращена к нижней стороне листовой пластинки. Первичная и вторичная флоэма, помимо различной мощности ситовидных элементов, внешне отличаются тем, что у первичной отсутствуют сердцевинные лучи. В состав флоэмы входят ситовидные элементы, паренхимные клетки, элементы сердцевинных лучей и механические элементы (табл. 7). Большинство клеток нормально функционирующей флоэмы живые. Отмирает лишь часть механических элементов. Собственно проводящую функцию осуществляют ситовидные элементы. Различают два их типа: ситовидные клетки и ситовидные трубки. Стенки ситовидных элементов содержат многочисленные мелкие сквозные канальцы, собранные группами в так называемые ситовидные поля. У ситовидных клеток, вытянутых в длину и имеющих заостренные концы, ситовидные поля располагаются главным образом на боковых стенках. Ситовидные клетки — основной проводящий элемент флоэмы у всех групп растений, исключая покрытосемянные. Клеток-спутниц у ситовидных клеток нет.
Ситовидные трубки покрытосемянных более совершенны. Они состоят из отдельных клеток — члеников, располагающихся один над другим (рис. 106). Длина отдельных члеников ситовидных трубок колеблется в пределах 150−300 мкм. Поперечник — составляет 20−30 мкм. Ситовидные поля этих члеников находятся плавным образом на их концах. Ситовидные поля двух расположенных один над другим члеников образуют ситовидную пластинку. Ситовидные поля представляют собой участки клеточной стенки, пронизанные многочисленными мелкими отверстиями, через которые с помощью цитоплазматических тяжей сообщаются протопласты соседних ситовидных элементов.
Членики ситовидных трубок формируются из вытянутых клеток прокамбия или камбия. При этом материнская клетка меристемы делится в продольном направлении и производит две клетки. Одна из них превращается в членик, другая — в клетку-спутницу. Наблюдается и поперечное деление клетки-спутницы с последующим образованием двух-трех подобных клеток, расположенных продольно одна над другой рядом с члеником. В клетках-спутницах вырабатываются различные ферменты, которые передаются в ситовидные трубки. При своем формировании членик ситовидной трубки имеет постенную цитоплазму, ядро и вакуоль. С началом функциональной деятельности он заметно вытягивается. На поперечных стенках появляется множество мелких отверстий-перфораций, образующих канальцы диаметром в несколько микрометров, через которые из членика в членик проходят цитоплазматические тяжи. На стенках канальцев откладывается полисахарид — каллоза, сужающий их просвет, но не прерывающий цитоплазматические тяжи.
По мере развития членика ситовидной трубки в протопласте образуются слизевые тельца. Ядро и лейкопласты, как правило, растворяются. Граница между цитоплазмой и вакуолью — тонопласт — исчезает, и все живое содержимое сливается в единую массу. При этом цитоплазма теряет полупроницаемость и становится вполне проницаемой для растворов органических и неорганических веществ. Слизевые тельца также теряют очертания, сливаются, образуя слизевый тяж и скопления около ситовидных пластинок. На этом формирование членика ситовидной трубки завершается. Длительность функционирования ситовидных трубок невелика. У кустарников и деревьев она продолжается не более 3−4 лет. По мере старения ситовидные трубки закупориваются каллозой и затем отмирают.
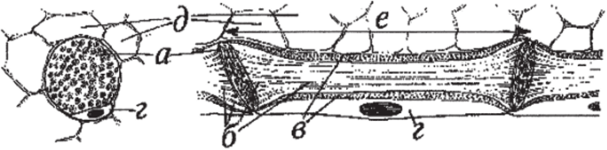
Рис. 106. Проводящие элементы флоэмы покрытосеменных на продольном (справа) и поперечном (слева) разрезах: а — ситовидная пластинка с первичными поровыми полями; б — плазмодссмы, проводящие растворы сахаров (фотоассимилятов); в — постенный слой цитоплазмы (в зрелом виде — без ядра); г — клетка-спутница (с ядром); д — лубяная паренхима; е — членик ситовидной трубки.
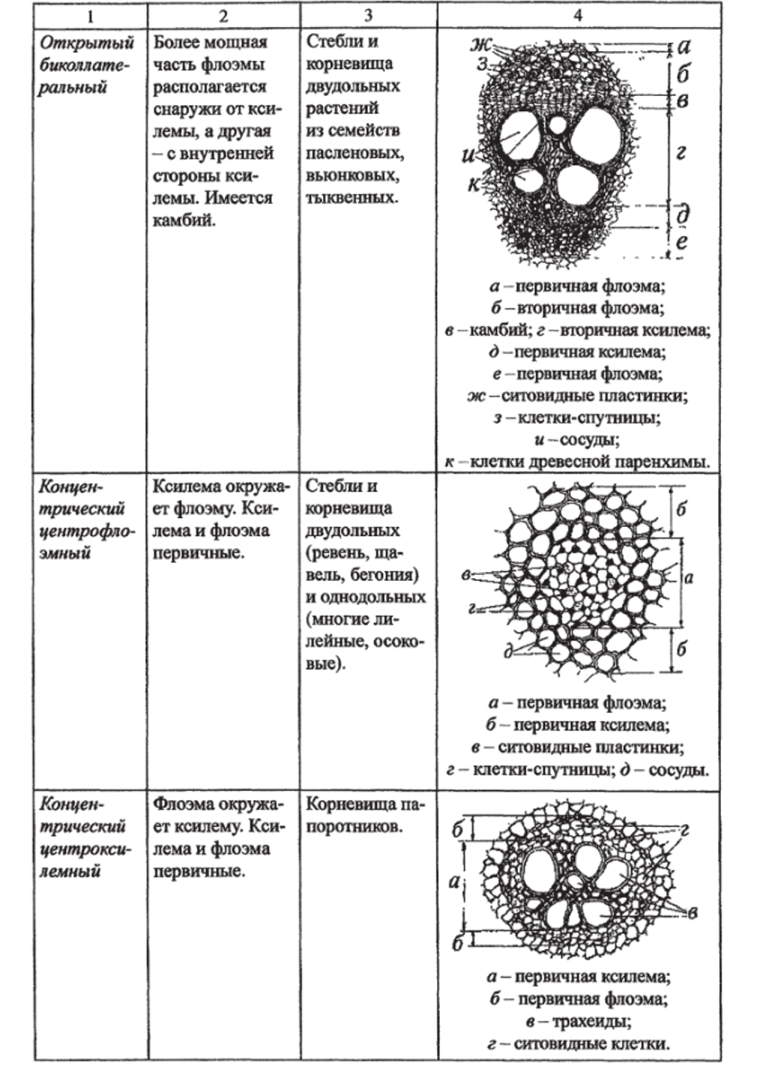
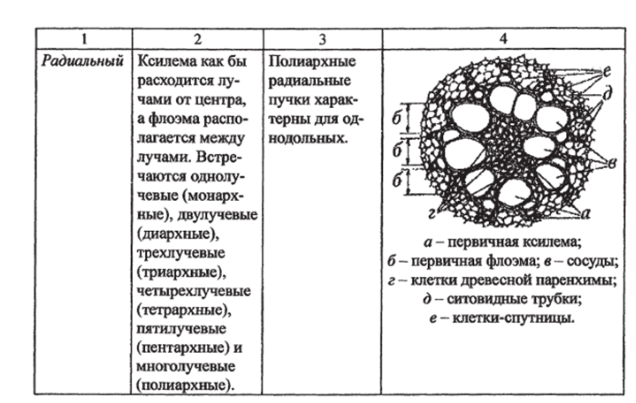
Типы проводящих пучков. Элементы проводящих пучков. Проводящая система семенных растений состоит из тяжей (комплекса ксилемы и флоэмы), которые называются проводящими пучками.
Таблица 8.
Типы проводящих пучков по способности к вторичному утолщению.
Закрытые. | Открытые. |
Прокамбий полностью дифференцируется в проводящие ткани и пучки далее не утолщаются. Характерны для однодольных растений. | Часть прокамбия сохраняется и превращается затем в камбий, пучки становятся способными к вторичному утолщению. Характерны для большинства двудольных и голосеменных. |
У разных растений проводящие пучки отличаются в зависимости от расположения в них ксилемы и флоэмы (табл. 9).
Таблица 9.
Типы проводящих пучков в зависимости от взаимного расположения флоэмы и ксилемы.
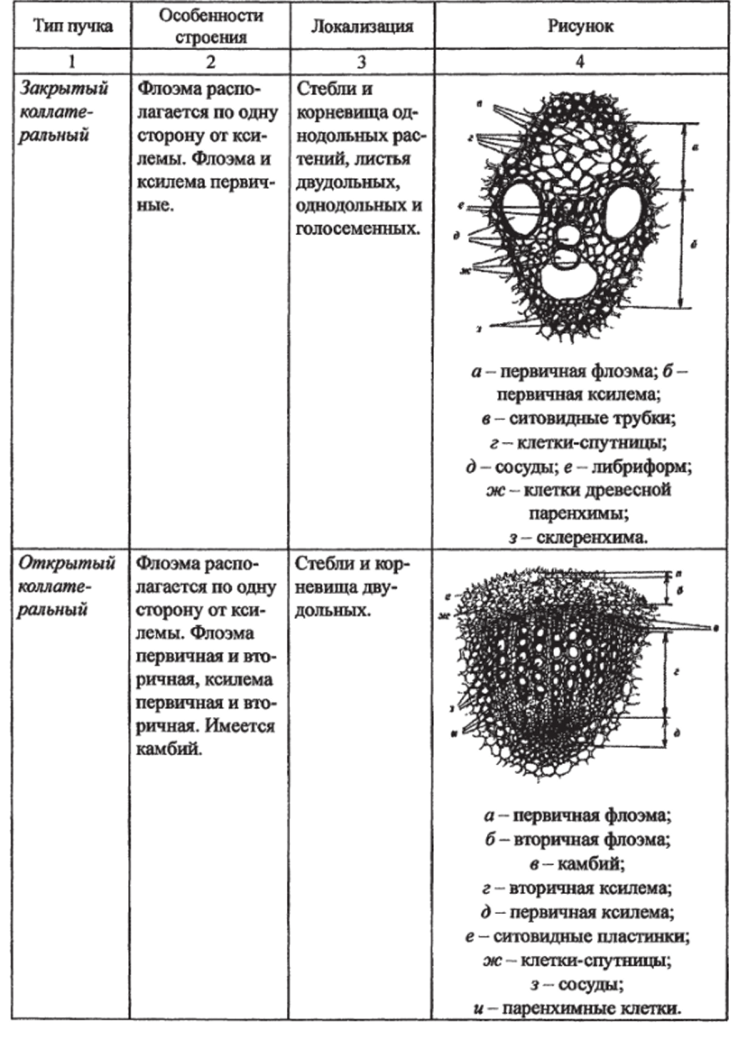
Основные ткани, их виды, строение, функции, локализация в органах растений. Основные ткани — это ткани, составляющие большую часть тела растения, относительно мало специализированные. Они занимают участки между другими постоянными тканями и присутствуют во всех вегетативных и репродуктивных органах. Они состоят из тонкостенных многоугольных живых паренхимных клеток, связанных с различными процессами жизнедеятельности растения. В зависимости от морфологических и физиологических особенностей выделяют следующие виды основных тканей:
- 1. Ассимиляционная (хлоренхима), у которой паренхимные клетки содержат хлоропласты, например, мезофилл листа. Хлоропласты в хлоренхиме располагаются не беспорядочно, а выстраиваются вдоль стенки (которая для увеличения поверхности может быть извилистой), не затеняя друг друга. Для мезофилла также характерен большой объем межклетников. Основные функции этой ткани — фотосинтез и ассимиляция органических веществ.
- 2. Запасающая с паренхимными клетками, способными накапливать продукты метаболизма в виде крахмальных зерен, алейроновых зерен и капелек масла (рис. 107). Наиболее выражена в специализированных запасающих органах: клубнях, луковицах и корневищах. В стеблях двудольных растений крахмал откладывается в эндодерме и является оберегаемым, то есть используется растением в неблагоприятных условиях, а сама эндодерма называется крахмалоносным слоем, или крахмалоносным влагалищем.
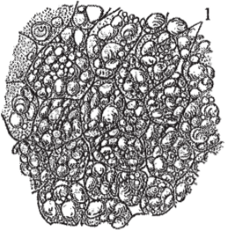
Рис. 107. Запасающая паренхима: 1 — крахмальные зёрна.
3. Воздухоносная ткань (аэренхима) имеет паренхимные клетки с крупными межклетниками, которые образуют хорошо развитую систему воздухоносных ходов, иногда проходящих непрерывно от листа до корня (рис. 108). Характерна для водных растений, у которых кроме функции аэрации может выполнять функции поддержания плавучести и повышения прочности.
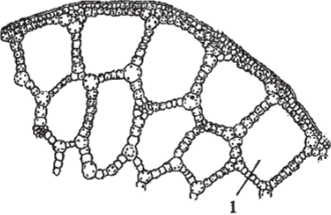
Рис. 108. Аэренхима: 1 - воздухоносный ход.
4. Водоносная, у которой паренхимные клетки имеют крупные вакуоли, содержащие слизь. Наличие слизи увеличивает способность клеток поглощать и удерживать воду. Характерна для растений-суккулентов (стеблевых и листовых).
Наружные и внутренние секреторные ткани, их строение и функции. Секреция — это комплекс процессов, связанных с отделением или изоляцией веществ от протопласта. Такими веществами могут быть избыточные ионы, удаляемые в виде солей, сахара, не утилизируемые или используемые частично продукты вторичного метаболизма (терпены, смолы и др.). Продукты секреции либо выделяются на поверхность растения или во внутренние вместилища, либо накапливаются в определенных частях клетки.
Секреторные структуры встречаются во всех органах растения и в зависимости от характера выделения подразделяются на внешние (экзогенные) и внутренние (эндогенные).
К внешним секреторным структурам принадлежат железистые волоски, железки, нектарники и гидатоды.
Железистые (головчатые) волоски, как правило, состоят из одно-многоклеточной ножки и одно-многоклеточной головки, в которой протекают процессы секреции (рис. 109).
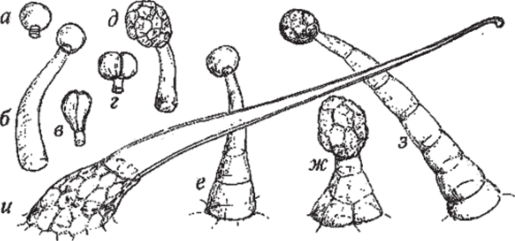
Рис. 109. Железистые (головчатые) волоски:
а, б — с одноклеточной головкой на одноклеточной ножке (короткой и длинной); в, г — с двухклеточной головкой (из кеглевидных или сферических клеток) на одноклеточной ножке; д — с многоклеточной головкой на одноклеточной ножке; е — с одноклеточной головкой на многоклеточной ножке; ж, з — с многоклеточной головкой на многоклеточной ножке; и — жгучий волосок крапивы.
Желёзки (в частности, эфирномасличные пельтатные желёзки губоцветных) также представляют собой, по сути, железистые волоски, но с очень короткой ножкой и многоклеточной головкой, покрытой общей кутикулой (рис. 110). Клетки головки продуцируют эфирное масло, выделяемое в суб кути кулярную полость, отчего последняя растягивается и после разрыва кутикулы, масло вытекает на поверхность растения. Железистые волоски (или пищеварительные желёзки) насекомоядных растений секретируют мукополисахариды, которые улавливают насекомых, и протеолитические ферменты, с помощью которых насекомые перевариваются. В специализированных солевых желёзках лебеды откладывается соль.
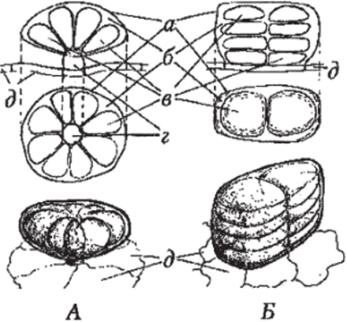
А Б
Рис. 110. Желёзки: А — губоцветных, Б — сложноцветных (вверху — ввд сбоку, в центре — сверху, внизу — в объёме): а — кутикула; б — секрет; в — секретирующие (выделительные) клетки; г — ножка; д — клетки эпидермы.
Нектарники секретируют жидкость, содержащую различные сахара. Различают флоральные нектарники, расположенные на цветках и экстрафлоральные нектарники, встречающиеся на вегетативных органах. Секреторная ткань нектарника может состоять только из эпидермы или из нескольких слоев нижележащих клеток, к которым близко подходит проводящая ткань. Причем, если в проводящей ткани преобладает флоэма, то нектар может иметь до 50% сахаров, а если ксилема, то их содержание может снижаться до 10%.
Гидатоды выделяют воду на поверхность листа и этот процесс называется гуттацией. Гидатоды обычно расположены по краю листа или на его верхушке, где поступающая вода из ксилемы может достигать поверхности через отверстия в эпидерме.
К внутренним секреторным структурам относятся секреторные клетки, секреторные полости и каналы, а также млечники.
Внутренние секреторные клетки (или идиобласты) заметно отличающиеся от остальных клеток имеют очень разнообразное содержимое. Например, во многих таких клетках накапливаются липофильные вещества, отчего клетки называют масляными (в частности, масляные клетки в корневищах аира болотного, в листьях и плодах многих лав ровых). В очень многих секреторных клетках содержимым являются танин и слизи (рис. 111).
Секреторные полости (или вместилища) и секреторные каналы отличаются от секреторных клеток тем, что они представляют собой пространства, которые возникают путём растворения клеток или отделения клеток друг от друга. В первом случае такие вместилища называются лизигенными, а во втором — схизогенными. В лизигенных вместилищах, по мере накопления секрета, клетки разрушаются, образуя полость. Схизогенные полости обычно выстланы неповрежденными (эпителиальными) клетками. К этому типу вместилищ относятся смоляные ходы хвойных, которые представляют собой длинные межклетники, выстланные продуцирующим смолу эпителием. Довольно часто секреторные полости и каналы возникают в результате повреждения растений (рис. 111).
Млечники представляют собой клетки или ряды клеток, содержащих латекс (млечный сок). По своему происхождению они могут быть простые (одиночные клетки) и сложные, возникающие из рядов клеток. Сложные млечники обычно называются членистыми, поскольку ряды клеток, составляющие их, соединяются путем растворения смежных стенок. Млечники обоих типов могут быть ветвистыми и неветвистыми. По цвету млечный сок может быть белым, как, например, у одуванчика, прозрачным, жёлтым (у чистотела), а по составу — весьма разнообразным. Так, частыми компонентами млечного сока являются терпены, в частности каучук, алкалоиды (у маковых), ферменты (у папайи) и другие группы соединений (рис. 111).
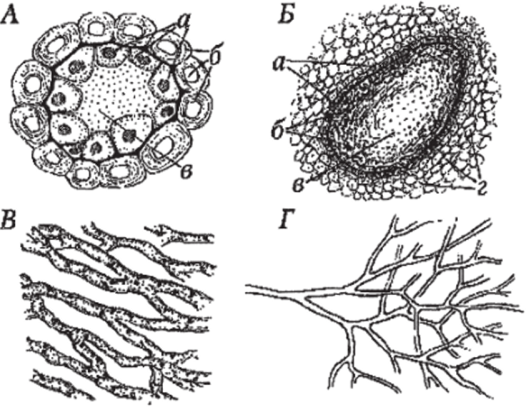
Рис. 111. Эндогенные секреторные структуры:
А — смоляной ход в поперечном разрезе: а живые секретирующие (выделительные) клетки с ядром; б — мертвые толстостенные клетки механической ткани; в — полость, заполненная секретом (смолой); Б — лизигенное вместилище: а — секретирующие клетки; б — лизированные клетки; в — полость вместилища, заполненная секретом; г — клетки основной ткани; В — членистые млечники в поперечном разрезе; Г — нечленистый млечник.
Тестовые задания для самоконтроля по теме «Растительные ткани (гистология)»
1. Обязательный элемент перидермы — это:
A) Устьица Б) Пробка.
B) Перицикл.
2. Камбий относится к ткани:
A) Покровной Б) Механической.
B) Образовательной.
3. Ситовидные трубки с клетками-спутницами относятся к элементам:
A) Ксилемы Б) Флоэмы.
B) Колленхимы.
4. Хлоренхима относится к:
A) Основной ткани Б) Выделительной ткани.
B) Покровной ткани.
5. Назовите вид основной ткани, в которой откладываются избыточные в данный период развития растений продукты метаболизма: белки, жиры, углеводы и др.:
A) Водоносная Б) Хлоренхима.
B) Запасающая.
6. Назовите первичную покровную ткань побега:
A) Склеренхима Б) Ксилема.
B) Эпидерма.
7. Механической тканью являются:
A) Склереиды Б) Трахеиды.
B) Рафиды.
8. Назовите эндогенные секреторные структуры:
A) Железки Б) Нектарники.
B) Млечники.
9. Назовите первичную покровную ткань корня:
A) Эпидерма Б) Эпиблема.
B) Колленхима.
10. Укажите вторичную латеральную меристему, формирующую вторичные ксилему и флоэму:
A) Прокамбий Б) Пери цикл.
B) Камбий.
11. Механическая ткань, состоящая из живых клеток, выполняющих арматурную функцию только в состоянии тургора и имеющих неравномерно утолщенные неодревесневающие первичные оболочки, подразделяющаяся на уголковую, пластинчатую и рыхлую:
A) Перидерма Б) Аэренхима.
B) Колленхима.
12. Полые трубки, состоящие из отдельных члеников, расположенных друг над другом и участвующих в проведении воды и растворенных в ней минеральных солей, это:
A) Сосуды Б) Млечники.
B) Ситовидные трубки с клеткам и-спутницами.
13. Назовите покровную ткань листьев:
A) Аэренхима Б) Колленхима.
B) Эпидерма.
14. Образовательная ткань, лежащая в основе перидермы — это:
A) Перицикл Б) Камбий.
B) Феллоген.
15. Механической тканью являются:
A) Эфирномасличные каналы Б) Трахеиды.
B) Лубяные волокна.
16. Назовите функции флоэмы:
A) Проведение воды и растворенных в ней минеральных веществ Б) Проведение органических веществ.
B) Газообмен и транспирация.
17. Назовите секреторные структуры, встречающиеся в экзокарпии плодов цитрусовых:
A) Смоляные ходы Б) Железки.
B) Лизигенные вместилища.