Общие черты строения и эмбрионального развития нервной системы позвоночных

Себе включает самостоятельные части, соответствующие основным группам современных рептилий. Анализ индексов энцефализации представителей вымерших групп рептилий, проведенный по эндокранам (динозавры, птерозавры, маммалоподобные рептилии), показал, что и их полигоны энцефализации не достигают уровня млекопитающих. Полигон амфибий расположен отчасти в зоне полигона рептилий. Обширный полигон… Читать ещё >
Общие черты строения и эмбрионального развития нервной системы позвоночных (реферат, курсовая, диплом, контрольная)
Нервная система всех позвоночных развивается из элементов наружного зародышевого листка — эктодермы. Этот процесс имеет определенные особенности у представителей разных групп, однако ему свойственны и общие для всех позвоночных закономерности.
Начальный этап развития нервной системы состоит в том, что на дорсальной стороне зародыша обособляется участок эктодермы — нервная пластинка, элементы которой интенсивно размножаются и дифференцируются, превращаясь в узкие цилиндрические нейроэпителиальные клетки, отличные от соседних клеток покровного эпителия. В результате интенсивного деления и неравномерного роста нейроэпителия происходит его инвагинация с последующим формированием нервной трубки. Замыкание нервной трубки начинается в середине зародыша, затем процесс распространяется к головному и хвостовому концам эмбриона, где некоторое время остаются незамкнутыми отверстия — передний и задний нейропоры.
Основной причиной формирования нервной пластинки и замыкания ее в нервную трубку является преобразование нейроэпителиальных клеток, связанное с изменением ориентации компонентов их цитоскелета. Нервная трубка (как полагают, под индуцирующим воздействием хорды) постепенно погружается в мезодерму зародыша и под влиянием мезодермальных сомитов разделяется на сегментарные участки — нейромеры. Сомиты располагаются по сторонам нервной пластинки и вдавливаются в нее, определяя конфигурацию будущих нейромеров (рис. 3).
У всех позвоночных развитие первой пары головных сомитов связано с формированием переднего мозга, но дальнейший процесс сегментации нервной системы идет по-разному у представителей разных групп позвоночных. Так, у хрящевых рыб и амфибий каждый последующий головной сомит соответствует по объему двум сомитам костистых рыб и амниот.
В дальнейшем головные сомиты сливаются и образуют три основных сегмента: премандибуляриый, мандибулярный и гиоидный. Границей головных сегментов служит область ушной капсулы, за которой формируются от 2 — 3 до 10 — 12 туловищных сегментов (в зависимости от группы позвоночных). Параллельно формируется система черепно-мозговых нервов. Каждый сегмент иннервируется определенными парами нервов: премандибуляриый — nervus tenninalis и глазодвигательным нервом (III); мандибулярный — тройничным (V) и блоковым (IV) нервами; гиоидный — отводящим (VI) и лицевым (VII) нервами. Следующие за головными два сегмента иннервируются соответственно языкоглоточным (IX) и блуждающим (X) нервами. Ростральные туловищные сомиты у высших позвоночных иннервируются системой добавочного нерва (XI), включающего в свой состав разное количество корешков в зависимости от числа туловищных сомитов. Подъязычный нерв (XII), иннервирующий гипобрахиальную мускулатуру, которая развивается из закладки туловищных сегментов, по своей функции аналогичен вентральным (соматомоторным) корешкам спинномозговых нервов, иннервирующих поперечнополосатую мускулатуру туловища и конечностей.
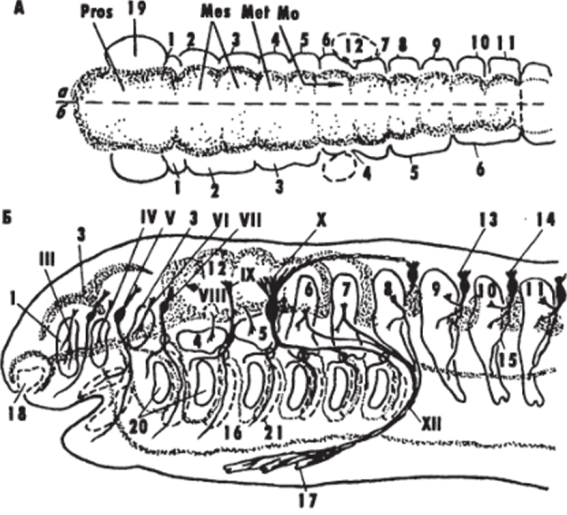
Рис. 3.
Схема развития мозговых пузырей (Л) и формирования головной части зародыша позвоночных (Б)
(Northcutt, 1979; Jacobson, 1988).
А: а. б — сомитомеры и мозговые пузыри костистых рыб и амниот (а), хрящевых рыб и амфибий (б). А, Б: 1 — 11 — сомитомеры (будущие туловищные сомиты); 12 — закладка ушной капсулы; спинальный ганглий; 14.15 — дорсальный (14) и вентральный (15) корешки спинномозгового нерва; 16 — полость целома; 17 — гипобрахиальная мускулатура; 18 — обонятельный орган, 19 — глазной пузырь; 20.21 — жаберные щели (20) и дуги (21); III — X, XII — черепномозговые нервы. Здесь и далее: остальные обозначения см. в списке условных сокращений Еще на стадии замыкания нейропоров начинается ростро-каудальная дифференцировка нервной трубки зародыша. Передний конец трубки значительно расширяется, боковые стенки утолщаются, образуя зачатки мозговых пузырей. Лежащий краниально пузырь образует первичный передний мозг, Prosencephalon, средний пузырь — первичный средний мозг, Mesencephalon, а из третьего пузыря, который переходит в закладку спинного мозга, развивается первичный задний мозг, Rhombencephalon.
Первый и третий мозговые пузыри вскоре посредством бороздсужений разделяются, образуя каждый по два вторичных мозговых пузыря. Эта стадия развития головного мозга получила название стадии пяти мозговых пузырей. Самым ростральным отделом нервной трубки становится конечный мозг, Telencephalon. Каудальнее располагается промежуточный мозг, Diencephalon, за которым следует средний мозг, Mesencephalon. Первичный задний мозг разделяется на вторичный задний мозг, Metencephalon, и продолговатый, Medulla oblongata.
Спинной мозг, Medulla spinalis, образуется из каудальных отделов нервной трубки. Он представляет собой часть ЦНС, в структуре которой наиболее отчетливо сохраняются черты эмбриональных стадий развития мозга позвоночных: трубчатый характер строения и сегментарность.
После формирования мозговых пузырей в нервной системе начинаются сложные процессы внутренней дифференцировки и роста. Уже на ранних этапах развития зародыша нервная трубка на значительном протяжении разделяется проходящей по вентрикулярной поверхности пограничной бороздой, sulcus limitans, на два отдела: дорсальный — крыловидная пластинка, и вентральный — базальная пластинка. Участки мозга, развивающиеся из крыловидной пластинки, содержат сенсорные ядра, из базальной — моторные и вегетативные. Ростральная часть нервной трубки не содержит базальной пластинки и целиком происходит из крыловидной. Отделы головного мозга, содержащие производные обеих пластинок, — средний, задний, продолговатый — часто объединяют названием «ствол мозга».
Разделение головного мозга на пять отделов (происходящих из мозговых пузырей) характерно для представителей всех групп позвоночных — от круглоротых до млекопитающих и человека, хотя структура и функциональная организация этих отделов претерпевают в эволюции позвоночных значительные изменения (рис. 4)[1].
Продолговатый мозг Medulla oblongata, является наиболее каудальным отделом головного мозга, плавно переходящим в спинной. Условной границей головного и спинного мозга считают ме<$то выхода первой пары спинномозговых нервов. Наиболее значительное сходство со спинным мозгом наблюдается в каудальных отделах продолговатого мозга и выражается в расположении серого и белого вещества, а также в центральном положении мозгового канала. Последний в ростральном направлении расширяется и превращается в ромбовидную или треугольную полость — IV мозговой желудочек, ventriculus quartus. Крыша его истончена и составлена лишь эпендимным эпителием, снаружи которого у высших позвоночных лежит сильно васкуляризованная мягкая оболочка мозга. У низших позвоночных серое вещество представляет собой тяжи, или столбы, нервных клеток, идущие вдоль продолговатого мозга, тогда как у высших позвоночных.

Рис 4.
Схема общей организации головного мозга низших (а) и высших (б) позвоночных
- (Platel, 1984)
- 1 — 4 — сосудистые сплетения конечного (/), промежуточного (2. 3), заднего (4) мозга; 5 — теменной орган; 6 — пинеальный орган (эпифиз); 7.8— верхние (7) и нижние (#) бугорки четверохолмия среднего мозга; vIII, vIV — желудочки промежуточного (vIII) и заднего (vIV) мозга; I. II — обонятельный (I) и зрительный (II) черепно-мозговые нервы намечается четкая тенденция к разделению этих тяжей на отдельные ядра.
Задний мозг, Metencephalon, у всех позвоночных представлен мозжечком, Cerebellum, форма, размеры и соотношение частей которого сильно варьируют у представителей разных групп позвоночных и в целом коррелируют со степенью совершенства их моторных реакций.
В вентральной и боковых стенках заднего мозга, слабо развитых у низших позвоночных, у млекопитающих формируется мощная система проводящих трактов и ядер, образующая самостоятельный отдел — мост, Pons. Полостью заднего мозга являются ростральные отделы IV мозгового желудочка.
Средний мозг, Mesencephalon, у низших позвоночных в дорсальных отделах сформирован крупными зрительными долями (tectum opticum), полости которых — зрительные желудочки — соединяются с общей полостью среднего мозга — водопроводом мозга, aqueductus cerebri. У млекопитающих на месте зрительных долей развиваются структуры четверохолмия, под которыми располагается зона покрышки. Вентральную часть среднего мозга у высших позвоночных занимают ножки мозга — система трактов, связывающих вышележащие отделы ЦНС с нижележащими.
Промежуточный мозг, Diencephalon, включает в себя лежащий дорсально эпиталамус, epithalamus, срединную часть — таламус, thalamus, и расположенный вентрально гипоталамус, hypothalamus. У высших выделяют несколько образований, формирующих субталамус, subthalamus, генез которого до конца не ясен.
Конечный мозг, Telencephalon, представляет собой наиболее сложный отдел головного мозга. Формирование его в эволюции различных групп позвоночных шло двумя принципиально разными путями, в связи с чем можно выделить два типа конечного мозга: инвертированный и эвертированный (рис. 5). Характерная черта строения инвертированного конечного мозга — наличие в полушариях латеральных, или боковых, желудочков. Такого типа мозг имеют круглоротые, хрящевые и двоякодышащие рыбы, амфибии, рептилии, птицы и млекопитающие. Полушария эвертированного типа лишены желудочков. Единственной группой позвоночных с таким типом конечного мозга являются лучеперые рыбы, Actinopterygii.
В конечном мозге выделяют лежащие дорсально паллиальные и расположенные вентрально субпаллиальные отделы. Первые представлены кортикальными формациями, вторые — ядрами конечного мозга. Подкорковые образования конечного мозга так же, как и кора, проходят сложный путь эволюции и имеют в своем составе филогенетически разные отделы: палеостриатум, архистриатум и неостриатум.
Таким образом, у всех позвоночных формируется сходный тип строения мозга. Однако следует еще раз подчеркнуть, что мозг позвоночных, претерпев длительную эволюцию, у современных их представителей обнаруживает удивительное разнообразие в степени развития одноименных отделов (рис. 6, 7).
Показано, что существует определенная зависимость между массой мозга, массой тела животного и уровнем развития его ЦНС. Эту зависимость выражают двумя коэффициентами: коэффициентом энцефализации (к) и коэффициентом аллометрии (а). Они вычисляются по формуле Е — кРа или чаще в логарифмической форме: log Е — log k + a log Р, где Е — масса мозга, Р — масса тела. Для каждого конкретного случая к влияет на угол наклона аллометрической кривой, определяя ее значение.
К настоящему времени указанные коэффициенты вычислены практически для представителей всех групп современных позвоночных. Так, для круглоротых коэффициент аллометрии составил 0,23 — 0,41; хрящевых рыб — 0,60; костистых рыб — 0,49; амфибий — 0,56 — 0,60; рептилий — 0,66; птиц — 0,36; млекопитающих — 0,63 — 0,67. Определение этих величин позволяет получить информацию о характере эволюционных преобразований мозга позвоночных и оценить уровень развития их нервной системы.
Значения коэффициентов аллометрии, рассчитанные для группы и для отдельных видов, могут существенно различаться. В пределах каждой группы позвоночных встречаются особи с высокими и низкими индексами энцефализации (I), рассчитываемыми по формуле I = ky/k0, где ку — коэффициент энцефализации данного.

Рис 5.
Два типа развития конечного мозга позвоночных а — ранняя стадия развития; б. в — инвертированный (б) и эвертированный (в) типы строения мозга. 1 — стриатум; 2 — септум; 3 — 5 — латеральная (3), дорсальная (4) и медиальная (5) зоны паллиума.
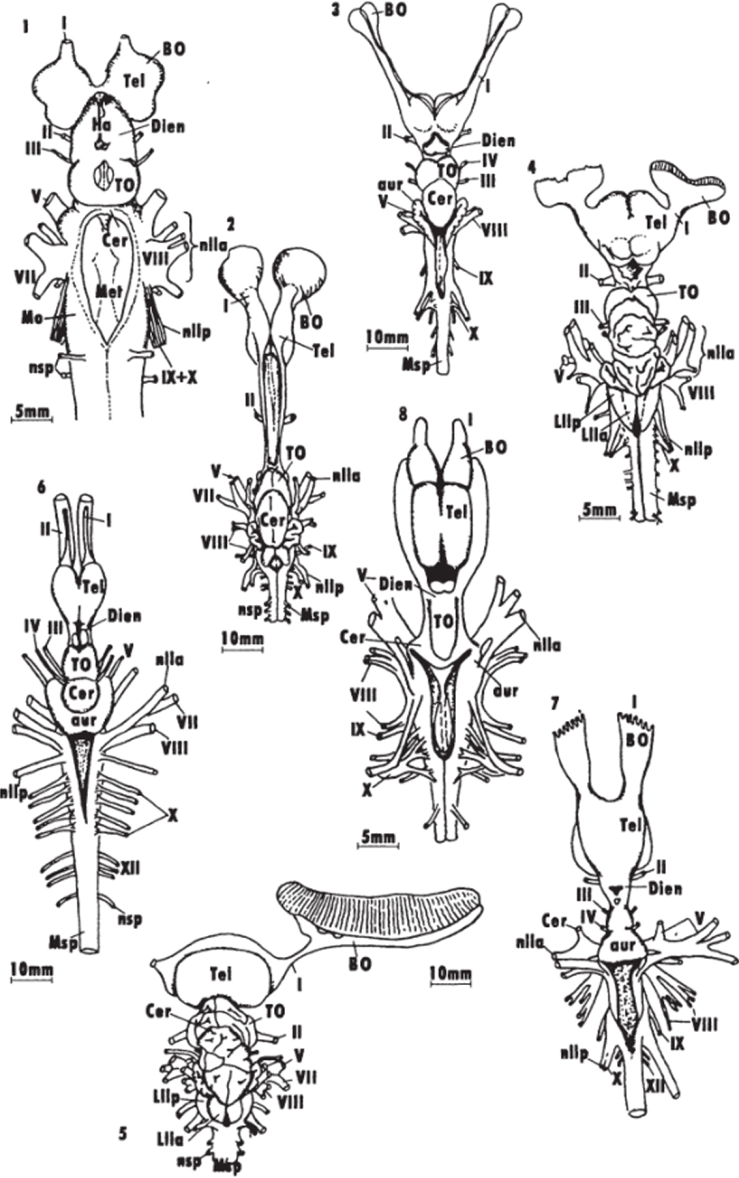
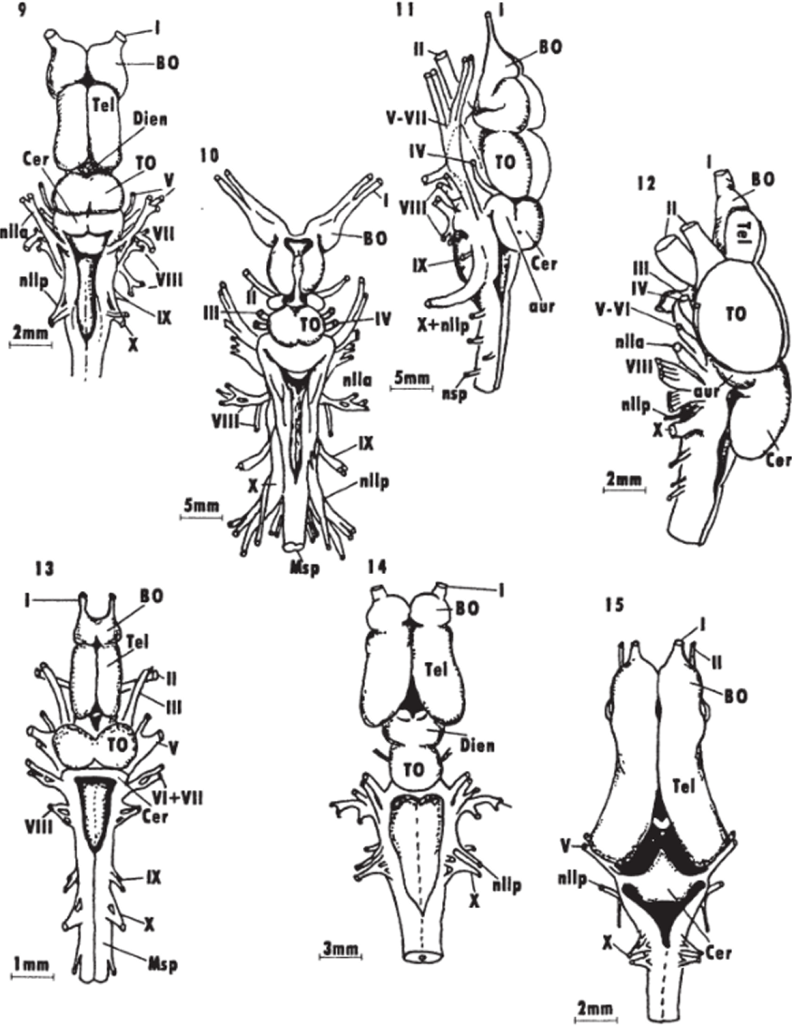
Ас. б Общий вид головного мозга представителей разных групп низших позвоночных (Northcutt, 1978, 1980, 1986; Northcutt, Puzdrowski.1988).
1 — - круглоротые (минога lchthyomyzan unicuspiв); 2 -5 — хрящевые рыбы: химеры Hydrolagus colliei (2), акулы: Notorynchus macululus (3), Musteluscanls (d). Sphyrna tyburo (5); 6 — # — capкоптсригии: латимерня iMtimrria chalumnar (G), двоякодышащие рыбы: Neoceratodus forsteri (7), l>ro1opterusannecteus (8); 9 — 12 — лучеперые рыбы: многопер Polipterus pal mas (fl), хрящевой ганоид Scapharhynchuspla torhynchus (10). костный ганоид Lepidusleus osseus (l 1), костистая рыба Satmo gaird fieri {12); 13—15 — амфибии: Rana cute$beiana (13), Amblystonia ligtinum (Id), Ichthyaphis glutinosus (15); I — XII — черепно-мозговые нервы животного, к0 — коэффициент энцефализации, принятый за точку отсчета. Например, индексы энцефализации ряда представителей приматов, рассчитанные относительно индекса энцефализации примитивных насекомоядных, составили: лемур — 429 — 629, бабуин — 924, макака-резус — 892, капуцин — 1038, горилла — 668, орангутан — 890, шимпанзе — 1131, человек — 2876.
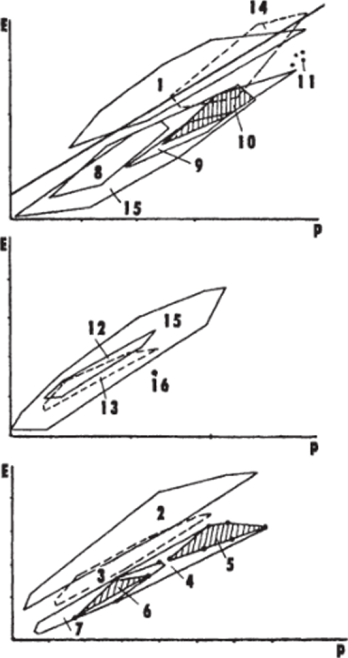
С учетом этого обстоятельства в сравнительной неврологии введено понятие «полигон энцефализации». Он представляет собой набор значений индексов энцефализации, определяемых отдельно для каждого вида и образующих некую геометрическую фигуру, ограниченную их крайними значениями. Такое построение позволило более тонко анализировать эволюционные изменения ЦНС в каждой конкретной группе позвоночных сравнительно с другими животными. Рассматривая полигоны энцефализации для основных групп современных позвоночных (рис. 8), можно отметить ряд закономерностей. Так, полигон энцефализации птиц четко отделен от полигона энцефализации рептилий, который сам по.
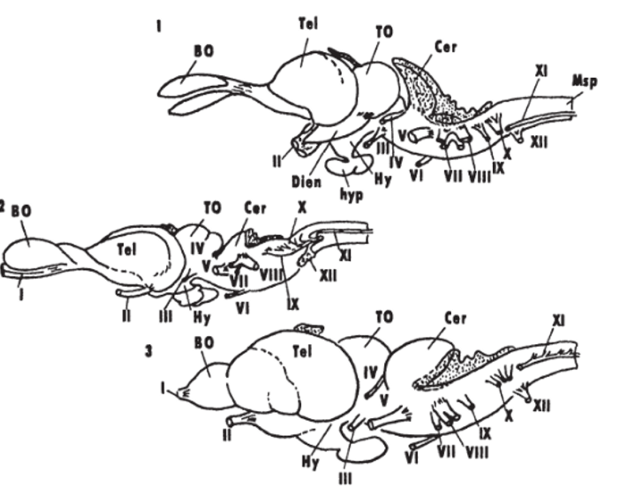
Рис 7.
Общий вид головного мозга рептилий (Bangma, 1983).
/ — ящерица Varanus exanthematlcus, 2— питов Pyton reglus, 3— черепаха Pseudemys crlspta elegans
себе включает самостоятельные части, соответствующие основным группам современных рептилий. Анализ индексов энцефализации представителей вымерших групп рептилий, проведенный по эндокранам (динозавры, птерозавры, маммалоподобные рептилии), показал, что и их полигоны энцефализации не достигают уровня млекопитающих. Полигон амфибий расположен отчасти в зоне полигона рептилий. Обширный полигон костистых рыб наглядно демонстрирует значительную эволюционную и адаптивную пластичность ЦНС у представителей многочисленных отрядов современных рыб, но и он, даже в своих высших значениях, нигде не пересекается с полигоном млекопитающих. Только хрящевые рыбы имеют индексы энцефализации, сравнимые с таковыми высших позвоночных — птиц и млекопитающих, что свидетельствует о высоком общем уровне организации их ЦНС.
По мнению нейроморфологов и нейрофизиологов, не всегда ясно адаптивное значение увеличения массы мозга, однако несомненно, что оно непосредственно не связано с явлениями пластичности и.
разнообразия функциональных связей в ЦНС. Таким образом, межвидовые и межгрупповые сравнения по массе, объему мозга или его отдельных частей, вычисление коэффициентов и индексов энцефализации и аллометрии могут служить только первым шагом в анализе сложных процессов морфо-функциональной эволюции нервной системы позвоночных.
Рис. 8
Полигоны индексов энцефализации основных групп позвоночных (современных и вымерших) (Platel, 1984)
/ — птицы; 2 — современные и 3 — древние млекопитающие; 4 — 11 — рептилии: современные и древние формы (4), динозавры (5). древние маммалоподобные (6), современные (7), ящерицы (5), змеи (9), черепахи (10), крокодилы (II); 12 — бесхвостые и 13 — хвостатые амфибий; 14 — хрящевые и 15 — костистые рыбы; /б — минога. Прямая линия — корреляционная кривая, рассчитанная для низших насекомоядных. Масштаб графиков не совпадает. По оси абсцисс — масса тела, по оси ординат — масса мозга.
- [1] Часто в составе мозга низших позвоночных два слабо дифференцированных отдела— промежуточный и конечный мозг — объединяют названием «передний мозг*.