Сцепление и кроссинговер

В другом эксперименте в качестве родителей были использованы мухи, обладающие теми же признаками, но в другом сочетании: мух с черным телом и нормальными крыльями скрещивали с мухами с нормальной окраской тела и зачаточными крыльями. Затем вновь провели 2 типа анализирующих скрещиваний: в первом использовали самцов Fu во втором — самок F{. В обоих случаях их скрещивали с двойным гомозиготным… Читать ещё >
Сцепление и кроссинговер (реферат, курсовая, диплом, контрольная)
В экспериментах с D. melanogaster Т. Х. Морган и его сотрудники обнаружили множество примеров сцепления генов и показали, что это сцепление как правило неполное.
Рассмотрим один из первых экспериментов Т. Х. Моргана по изучению сцепленного наследования.
У дрозофилы известны мутантные популяции, отличающиеся от мух дикого типа черной окраской тела. Это признак рецессивный по отношению к признаку нормальной серой окраски. Ген, контролирующий черную окраску тела, называется black и обозначается Ь. Его доминантный аллель — Ь*.
Существует также рецессивный геи vestigial (vg), который в гомозиготном состоянии приводит к недоразвитию крыльев (зачаточные крылья). Его доминантный аллель (vg+) контролирует нормальное развитие крыльев.
При скрещивании мух $ bb vg vg х b* b* vg* vg' в /^были получены особи, дигетерозиготные по этим генам. Все они были нормальными по обоим признакам в соответствии с правилом доминирования и законом единообразия F,. Далее были проведены два типа анализирующих скрещиваний. В первом из них брали самцов F и скрещивали с гомозиготными самками bb vg vg, а во втором — девственных самок, отобранных в F, скрещивали с самцами bb vg vg (рис. 4.1, а). Результаты этих анализирующих скрещиваний оказались неодинаковыми. В F (потомство от анализирующего скрещивания) в первом случае были получены мухи только двух типов независимо от пола: 50 % мух имели черное тело и зачаточные крылья, 50 % были нормальными по обоим признакам. Учитывая, что расщепление в анализирующем скрещивании отражает соотношение типов гамет, продуцируемых особями F{, следует заключить, что самцы F, использованные при первом скрещивании, формировали гаметы только двух типов — с родительскими сочетаниями аллелей b vg и F vg*. Следовательно, в 100% случаев образовывались гаметы только с родительскими сочетаниями исследованных генов.
При втором скрещивании в F появились все возможные четыре типа потомков, следовательно, самки Fi давали четыре типа гамет: b vg, b* vg, b vg* и b*vg*. В Fa от этого скрещивания четыре типа гамет образовались в разном количестве (рис. 4.1, б). Независимо от пола мухи распределились следующим образом: 41,5% черных с зачаточными крыльями; 41,5 % нормальных по окраске и с нормальными крыльями; 8,5 % черных с нормальными крыльями; 8,5% нормальных по окраске с зачаточными крыльями. Таким образом, родительские сочетания b vg и b* vg* образовались в 83% случаев, а новые комбинации — рекомбинантные сочетания b vg* и b* vg — в 17% случаев.

Рис. 4.2. Родительские и рекомбинантные сочетания генов.
В другом эксперименте в качестве родителей были использованы мухи, обладающие теми же признаками, но в другом сочетании: мух с черным телом и нормальными крыльями скрещивали с мухами с нормальной окраской тела и зачаточными крыльями. Затем вновь провели 2 типа анализирующих скрещиваний: в первом использовали самцов Fu во втором — самок F{. В обоих случаях их скрещивали с двойным гомозиготным рецессивом bb vg vg, и вновь получили такие же результаты в отношении родительских и рекомбинантных сочетаний признаков в Fa. Если из брали самцов, то наблюдали только родительские комбинации признаков, а если из Fx брали самок, то появлялись родительские (83%) и рекомбинантные (17%) сочетания признаков в тех же соотношениях, что и в первом эксперименте.
В обоих эксперимешдх наблюдается полное сцепление генов bwvg, если для анализирующего скрещивания берут самцов F. Если же для анализирующего скрещивания используют самок F9 то сцепление частичное.
Т.Х. Морган дал следующее объяснение этим результатам. В первом эксперименте гены b и vg находятся в одной хромосоме, т. е. дигетерозиготные особи F несут в одном гомологе аллели b и vg, а в другом гомологе — Ь* и vg*; во втором эксперименте b и vg* в одном, а Ь* и vg — в другом гомологе.
У самцов дрозофилы кроссинговер вообще нс происходит, поэтому гены, локализованные в одной хромосоме (или, говоря более строго, в одной паре хромосом), обнаруживают абсолютное сцепление, если при скрещивании используют дигетерозиготных самцов. В мейозе у дигетерозиготных самок дрозофилы F возможен обмен гомологичными участками гомологичных хромосом между локусами, в которых находятся гены b и vg. Такие обмены, или кроссинговер (от англ, crossingover — перекрест), приводят к новому рекомбинантному сочетанию аллелей генов b и vg в гомологичных хромосомах, которые затем расходятся к разным полюсам. Эти обмены происходят с вероятностью 17% и в итоге дают два класса реципрокных рекомбинантных сочетаний, или рекомбинантов с равной вероятностью — по 8,5 %.
Сходным образом объясняется и результат, полученный ранее У. Бэтсоном и Р. Пеннетом: гены, контролирующие окраску цветков (р) и форму пыльцевого зерна (/) у душистого горошка, локализованы в одной паре гомологичных хромосом, и между ними возможен кроссинговер.
Сотрудник Т. Х. Моргана А. Стсртсвант предположил, что частота кроссинговера на участке между генами, локализованными в одной хромосоме, может служить мерой расстояния, на котором они находятся друг от друга. Тогда можно использовать частоту кроссинговера для того, чтобы определять взаимное расположение генов и расстояние между ними.
В качестве подтверждения справедливости этого положения можно в общем виде рассмотреть результаты тригибридного скрещивания, в котором родительские формы дрозофилы различаются по уже известным нам генам b и vg, а также дополнительно по гену рг, проявляющему сцепление с Ь и vg. Рецессивный аллель pr {purple — пурпурный) в гомозиготном состоянии обусловливает ярко-красную окраску глаз, нормальная доминантный аллель рг+ — темно-красный цвет глаз.
При анализирующем скрещивании потомки расщепляются на 8 классов: 2 класса нерекомбинантных (I и II) и 6 классов потомков, рекомбинантных по всем генам (Ш-VIII).
Далее необходимо определить частоту кроссинговера между всеми тремя генами попарно. Для этого суммируют количество всех мух, рекомбинантных по генам Ъ и рг. классы Ш, IV, VII, VIII. Полученное число делят на общее число исследованных потомков в Fa. Аналогично определяют частоту рекомбинации (кроссинговера) между рг и vg (при этом суммируют классы V, VI, VII, VIII) и частоту рекомбинации между b и vg (суммируют классы Ш, IV, V, VI).
Экспериментально установленные частоты рекомбинации между тремя генами попарно можно представить следующим образом:
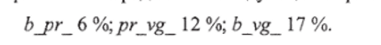
На основании этих данных, пользуясь правилом аддитивности, можно расположить 3 гена в линейной последовательности. Наиболее удалены друг от друга гены b и vg, а между ними локализован ген рг. Сумма частот его рекомбинации с генами b и vg приблизительно равна частоте рекомбинации между Ь и vg. Таким образом, строится простейшая карта группы сцепления.
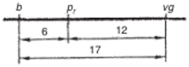
Рис. 4.3. Карта группы сцепления.
В строгом смысле группой сцепления называют группу генов, проявляющих сцепленное наследование. Поскольку известно, что такое наследование отражает локализацию генов в одной хромосоме, обычно под группой сцепления понимают группу генов, расположенных в одной хромосоме.
Гены, расположенные в одной хромосоме, не всегда обнаруживают сцепление. Генетичсское расстояние, на котором кроссинговер происходит с вероятностью I %, измеряется в сантиморганах (сМ) — эта единица измерения, названная в честь Т. Х. Моргана.