Предпосылки молекулярной модели кроссинговера
Следует напомнить так называемую высокую отрицательную интерференцию. Явление, которое трактовали как стимулирование одного реципрокного обмена другим, также наблюдается на очень коротких генетических расстояниях, например в IV хромосоме D. melanogaster, имеющей рекомбинационную длину всего 3,0%. Появление в полигибридных скрещиваниях множественно-рекомбинантных сегрегантов по очень тесно… Читать ещё >
Предпосылки молекулярной модели кроссинговера (реферат, курсовая, диплом, контрольная)
Как уже отмечалось, попытка объяснить появление кроссоверных классов только на основе гипотезы конверсии, т. е. взаимопревращения аллелей в дигетерозиготе, оказалась несостоятельной. Реальность реципрокных обменов была доказана при цитогенетическом изучении кроссинговера. Гипотезу конверсии, казалось бы, опровергают данные тетрадного анализа — регулярное расщепление (2: 2) для каждого из исследуемых генов. Действительно, эта иллюстрация правила чистоты гамет опирается на наиболее частые варианты тетрад.
Сенсационным стало открытие К. К. Линдегрена (1949), описавшего отклонения от нормального (2: 2) расщепления в тетрадах диплоидного гибрида дрожжей Sacch. cerevisiae, гетерозиготного по ADE2: adel. Наряду с тетрадами 2 ADE2: 2 ade2 изредка встречались тетрады с соотношением 3 ADE2: ade2 и 1 ADE2: 3 ade2.
В дальнейшем было показано, что у дрожжей конверсия по различным генам — обычное явление, встречающееся с частотой около 1%. Как правило, наблюдается равенство частот тетрад 3А: 1а и А: За. Конверсия была описана и у других грибов, например Neurospora crassa, Sordaria fimicola. При этом у них были получены не только октады с соотношением 6А: 2а или 2А: 6а, аналогичные тетрадам дрожжей с расщеплением ЗА: а и А: За, но и октады с соотношением 5: 3 (и 3: 5), поначалу казавшиеся не соответствующими расщеплению по закону Г. Менделя. Появление таких соотношений можно объяснить, только предположив, что у некоторых гаплоидных продуктов мейоза происходит расщепление в последующем митотическом делении — так называемое постмейотическое расщепление.
Если конверсию изучают в полигетерозиготах по ряду сцепленных маркеров, можно наблюдать корреляцию между конверсией по какомулибо маркеру и реципрокной рекомбинацией по фланговым маркерам, т. е. по маркерам, расположенным слева и справа от участка конверсии. С. Фогель, Р. Мортимер и Д. Херст изучили 11 023 тетрады дрожжей, среди которых обнаружили 907 случаев конверсии {конвертантных тетрад), и убедились, что в 445 из них (49,1%) одновременно с конверсией произошла реципрокиая рекомбинация (кроссинговер) по фланговым маркерам, т. е. практически в половине случаев конверсия сопровождалась кроссинговером. Исследователи получили доказательство того, что конверсия не приводит к появлению новых аллелей, а только нарушает соотношение родительских аллелей при расщеплении некоторых тетрад. Это заставило предположить, что конверсия происходит вследствие копирования одного аллеля за счет другого. Здесь уместно вспомнить гипотезу Дж. Беллинга, возрожденную в 1950;х гг. А. Херши и Дж. Ледербергом, о выборочном копировании, или копировании со сменой матриц, применительно к репликации и рекомбинации геномов бактериофагов и бактерий. Согласно этой гипотезе репликация частично может происходить по матрице одной, а частично — по матрице другой (гомологичной) молекулы ДНК (рис. 6.6). По-видимому, данный механизм возможен только для рекомбинации на очень коротких участках, иначе это будет противоречить полуконсервативному механизм)' репликации ДНК.
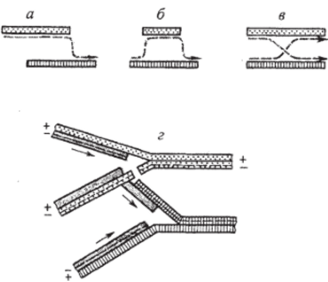
Рис. 6.6. Различные схемы механизма выборочного копирования: а — образование рекомбинантной хромосомы по гипотезе частичных реплик; б — выборочное копирование части хромосомы; в- реципрокное переключение реплик, обеспечивающее кроссинговер; г — разрыв и копирование; а, б, в: сплошные линии — двунитевые молекулы ДНК, пунктирные — их копии при допущении консервативной репликации; г: антипараллельные нити ДНК (+ и -) изображены сплошными и пунктирными линиями. Стрелки — направление репликации На участие матричного процесса (возможно, репликации) в ходе конверсии указывают и данные по совместной конверсии сразу двух тесно сцепленных мутаций. Например, в тетрадном анализе дигетерозиготы дрожжей 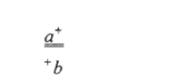
могут быть получены тетрады как с аномальным расщеплением по каждому из маркеров:
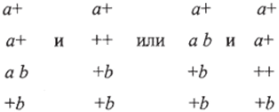
так и с аномальными расщеплениями по обоим маркерам:
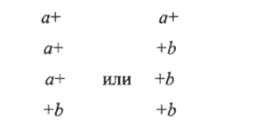
Это явление получило название совместной конверсии, или коконверсии. Могут быть получены и тетрады — результат реципрокной рекомбинации:
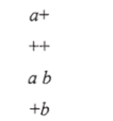
Как правило случаи коконверсии встречаются на коротких расстояниях между а и b (около 1 % рекомбинации), причем чем ближе располагаются а и by тем чаще происходит коконверсия и тем реже — реципрокная рекомбинация. Существование коконверсии показывает, что конверсия захватывает некоторый участок генетического материала.
Следует напомнить так называемую высокую отрицательную интерференцию. Явление, которое трактовали как стимулирование одного реципрокного обмена другим, также наблюдается на очень коротких генетических расстояниях, например в IV хромосоме D. melanogaster, имеющей рекомбинационную длину всего 3,0%. Появление в полигибридных скрещиваниях множественно-рекомбинантных сегрегантов по очень тесно сцепленным маркерам дает величину коинциденции с" 1, и интерференция (/= 1 — с) в таком случае приобретает отрицательное значение. Подобные рассуждения справедливы только в случаях с реципрокной рекомбинацией — кроссинговером. Кажущееся совпадение нескольких рекомбинационных событий на коротких расстояниях — результат конверсии, что обнаруживается только при тетрадном анализе. Например, при расщеплении тригетерозиготы.
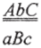
могут появиться тетрады:

с конверсией по среднему маркеру В/b. При рассмотрении случайной выборки гамет при генетическом анализе высших эукариот, например при анализирующем скрещивании, появление потомков АВС и abc создает впечатление неслучайного совпадения обменов на участках A-В и В-С при тесном сцеплении всех маркеров и редком появлении одиночных рекомбинантов на участке А-С.
В действительности это результат конверсии, или нереципрокной рекомбинации, а так называемой высокой отрицательной интерференции не существует.