Транспозируемые (мигрирующие) генетические элементы

Полагают, что специфические концевые повторы /S-элементов и транспозонов играют ключевую роль в процессах транспозиции, поскольку они распознаются ферментами, осуществляющими сайт-специфическую рекомбинацию. Вместе с тем в случае ряда транспозонов получены экспериментальные данные, свидетельствующие о том, что при транспозиции может работать механизм, не связанный с процессом полного «вырезания… Читать ещё >
Транспозируемые (мигрирующие) генетические элементы (реферат, курсовая, диплом, контрольная)
Согласно представлениям, сложившимся в рамках классической (формальной) генетики, геномы различных организмов характеризуются значительной стабильностью и подвержены лишь очень медленным эволюционным преобразованиям. Эти представления существенно изменились в связи с открытием транспозируемых (мигрирующих) генетических элементов (фрагментов ДНК), которые могут перемещаться как в пределах одного генома, так и из одного генома в другой. Способность к транспозиции закодирована в нуклеотидных последовательностях самих мигрирующих фрагментов ДНК и связана с возможностью их включения (интеграции) в различные места (сайты) хромосом и внехромосомных молекул ДНК и выхода («вырезания») из этих сайтов. Такие мобильные структуры были обнаружены в геномах как прокариот, так и эукариот.
У бактерий транспозируемые генетические элементы представлены сегментами ДНК двух типов: инсерционными (вставочными) последовательностями, или /5-элементами (от англ, insertion sequences — вставочные последовательности), и транспозонами. Те и другие могут перемещаться в границах одной генетической структуры (бактериальной хромосомы, плазмиды) либо из одной структуры в другую (например, из хромосомы в плазмиду, из плазмиды в хромосому, из одной плазмиды в другую, из плазмиды бактерии одного вида в плазмиду либо хромосому бактерии другого вида и т. д.).
/5-элементы — это небольшие участки ДНК, размеры которых обычно варьируют в пределах от 760 до 2 000 пар нуклеотидов. На концах (флангах) этих участков имеются одинаковые либо очень сходные короткие нуклеотидные последовательности (20−40 пар нуклеотидов), расположенные в обратном порядке (инвертированные повторы). Считают, что концевые повторы (фланкирующие последовательности) играют важну ю роль в процессах перемещения транспозируемых элементов, хотя точный механизм таких перемещений недостаточно изучен.
Разные /5-элемснты являются типичными «обитателями» бактерий и обычно выявляются в количествах, превышающих одну' копию на клетку (табл. 5.6). Наличие идентичных /5-элементов в разных бактериальных структурах (хромосомах, плазмидах) создает основу для их взаимодействия по механизму сайт-специфической рекомбинации, что может приводить к объединению (коинтеграции) таких структур либо к генетическому обмену между ними. В отличие от генетической рекомбинации общего типа, требующей наличия значительных областей гомологии ДНК у двух взаимодействующих родственных структур, сайт-специфическая рекомбинация основана на том, что соответствующий фермент узнает небольшие специфические последовательности в двух местах (сайтах) одной либо разных молекул ДНК, где и происходит процесс рекомбинации. Следовательно, такая рекомбинация может приводить к генетическому обмену между неродственными (негомологичными) структурами (хромосомами, плазмидами).
Таблица 5.6
Содержание инсерционных последовательностей в хромосомном геноме Е. соН
Название. | Число копий в клетке. | Размеры одной копии. |
последовательности. | на хромосому. | (в парах нуклеотидов). |
/51. | 5−8. | |
/52. | 1 327. | |
/53. | 1 400. | |
/54. | 1−2. | 1 400. |
Транспозоны являются более сложными образованиями с размерами порядка 2 000−20 000 пар нуклеотидов, в срединной части которых находятся те или иные функционально значимые гены, а концевые участки содержат /S'-элементы либо фланкированы иными длинными инвертированными или одинаковыми (прямыми) нуклеотидными повторами (800−1 500 пар нуклеотидов). В составе некоторых транспозонов обнаружены гены, кодирующие синтез ферментов, названных транспозазами и резолвазалш, которые считаются ответственными за процессы транспозирования. Вместе с тем транспозоны могут содержать гены, не имеющие отношения к транспозиции, но играющие важную роль в выживании бактерий при неблагоприятных условиях среды обитания (например, гены устойчивости к антибиотикам и другим антибактериальным препаратам). Такие гены перемещаются вместе со всем транспозоном из одной бактериальной плазмиды в другую, из плазмиды в хромосому ит.д.
Полагают, что специфические концевые повторы /S-элементов и транспозонов играют ключевую роль в процессах транспозиции, поскольку они распознаются ферментами, осуществляющими сайт-специфическую рекомбинацию. Вместе с тем в случае ряда транспозонов получены экспериментальные данные, свидетельствующие о том, что при транспозиции может работать механизм, не связанный с процессом полного «вырезания» всей структуры из одного сайта и ее перемещения в другой сайт (сайтмишень). При этом происходит удвоение (репликация) генетического элемента, одна копия которого остается в исходном сайте, а другая перемещается в новый сайт-мишень. Иными словами, такая транспозиция состоит в увеличении числа копий соответствующего транспозона. Если же все-таки происходит исключение («вырезание») транспозона, то этот процесс не всегда бывает достаточно точным и может приводить к сохранению в структуре ДНК бывшей мишени тех или иных остатков последовательностей транспозируемого элемента. Известно также, что встраивающийся транспозон либо /S-элемснт фланкируется короткими повторами ДНК хозяина (4−12 пар нуклеотидов). Есть основания считать, что транспозируемые элементы играют важную роль в процессах эволюции микроорганизмов, обеспечивая специфическую форму их внутривидового и межвидового генетического обмена (так называемый горизонтальный перенос генов). Известно также, что при интеграции мобильных структур в соответствующие сайты хромосом и плазмид и при их «вырезании» в этих и смежных сайтах могут происходить мутационные перестройки генетического материала (инверсии, делеции, транслокации и др.), т. е. транспозируемые элементы в этом случае выступают в роли биологических мутагенов.
Мигрирующие генетические элементы эукариот, впервые обнаруженные в 1951 г. у высших растений (кукурузы), были затем выявлены и у других организмов. Содержание некоторых из этих элементов в эукариотическом геноме может быть весьма значительным, т. е. они являются многокопийными нуклеотидными последовательностями ДНК. Вместе с тем функциональное значение таких последовательностей остается во многом неясным.
Мобильные структуры небольших размеров, содержащие концевые повторы и наиболее сходные с транспозонами бактерий, были обнаружены в различных участках геномов дрожжей и дрозофилы. К числу наиболее изученных мигрирующих элементов дрозофилы относится группа близких по строению нуклеотидных последовательностей, обозначенных общим термином copia, которые в совокупности составляют значительную часть геномной ДНК этих организмов.
В различных участках генома человека обнаруживается многократно повторяющаяся (около 500 тыс. копий на гаплоидный геном) нуклеотидная последовательность, состоящая из 300 пар нуклеотидов, которая получила название Л/ы-последовательности. Поскольку концевые участки этой последовательности фланкированы короткими прямыми повторами (7−20 пар нуклеотидов), определяющими ее сходство с известными транспозонами, то полагают, что она также относится к категории мобильных элементов эукариот. Сходные Л/ы-подобные последовательности выявлены и в геномах различных млекопитающих.
В качестве источника мобильных структур прокариотического и эукариотического генома можно рассматривать также геномную ДНК различных вирусов, которые способны интегрироваться в те или иные участки ДНК бактерий и эукариот и некоторое время существовать там в форме провируса, т. е. как составная часть генетической структуры клеткихозяина. Примером является интеграция бактериофага X в хромосомную структуру клетки Е. coli с помощью механизма сайт-специфической рекомбинации (рис. 5.31). Эта особенность касается и РНК-содержащих ретровирусов, ДНК которых, синтезированная на основе рассмотренного выше механизма обратной транскрипции, включается в хромосомную структуру эукариотической клетки и затем может длительно реплицироваться в ее составе, передаваясь от материнской клетки к дочерним в процессе митотического деления. При последующем неправильном (неточном) «вырезании» из генома хозяина вирус оставляет в нем те или иные фрагменты собственного генома и приобретает отдельные участки генетического материала хозяина.
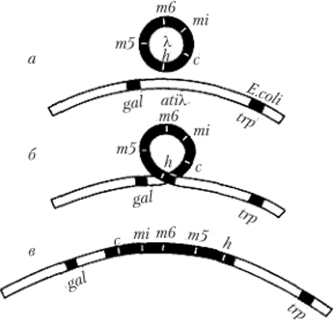
Рис. 5.31. Интеграция бактериофага X в хромосомный сегмент клетки Е. coli между локусами генов gal (утилизации галактозы) и trp (синтез триптофана): а — кольцевая хромосома фага X (m5, m6, mi, c, h- отдельные гены фаговой ДИК) и фрагмент кольцевой хромосомы Е. coli (att Х-сайт гомологии с фаговой ДНК); б — сайт-специфическая рекомбинация, связанная с разрывами хромосомы фага между локусами Лиси бактериальной хромосомы между локу сами gal и trp; в — бактериальная хромосома, содержащая ДНК фага X в форме профага Можно полагать, что результатом указанных событий являются генетические элементы клеток животных и человека, названные резроэлеменгами (рстротранспозонами), способные к перемещениям благодаря наличию у них коротких концевых повторов. Такие ретротранспозоны могут, в частности, кодировать белки, необходимые для осуществления в эукариотической клетке процесса обратной транскрипции PHК-содержащих ретровирусов, которые рассматриваются в качестве причины возникновения злокачественных опухолей (онкологических заболеваний) и синдрома приобретенного иммунодефицита (СПИДа). Так, получены экспериментальные данные, свидетельствующие о том, что некоторые онкогены клеток человека, связанные с появлением злокачественных новообразований, имеют структурную гомологию с РНК ретровирусов.
Значение транспозируемых элементов эукариот связывают прежде всего с их мутагенным действием, а также с вероятным участием в процессах генетической регуляции, поскольку, интегрируясь по соседству с некоторыми генами, они могут оказывать существенное влияние на их экспрессию. Становится очевидной также роль ретротранспозонов, представленных нуклеотидными последовательностями некоторых ретровирусов, в развитии ряда заболеваний животных и человека (лейкозы птиц и млекопитающих, злокачественные новообразования, СПИД и др.). Следует еще раз подчеркнуть вероятную роль мигрирующих элементов как одного из факторов эволюции организмов.
Таким образом, экспериментальные достижения, которые привели к расшифровке структуры генетического кода и установлению механизмов его реализации, не только открыли новые возможности понимания живой природы, но и обеспечили решение ряда практических задач на основе манипуляций с молекулами ДНК из организмов разных видов (успехи в области генетической инженерии, биотехнологии и др.). Эти достижения послужили также основой для разработки новых направлений в медицинской и клинической генетике, связанных с диагностикой и лечением наследственной патологии человека (генная диагностика, генная терапия и др.).