Зрительная система.
Нервная система: анатомия, физиология, нейрофармакология

Видимое изображение фоторецепторы «описывают» как меняющуюся в пространстве и во времени совокупность красных, зеленых, синих и серых точек. Данная информация разделяется в результате дополнительной обработки клетками сетчатки на несколько параллельных потоков, которые в виде нервных импульсов поступают в ГМ по зрительному нерву. Работу сетчатки можно сравнить с работой матрицы фотоаппарата или… Читать ещё >
Зрительная система. Нервная система: анатомия, физиология, нейрофармакология (реферат, курсовая, диплом, контрольная)
Зрение представляет собой процесс получения информации о внешнем мире с помощью восприятия электромагнитных волн с определенной длиной. Диапазон видимого света у человека составляет 400—750 нм. Самые короткие волны субъективно воспринимаются как фиолетовые, самые длинные — как красные.
Глаз человека имеет шарообразную форму, за что получил название глазного яблока. Три пары мышц, иннервируемых глазодвигательными нервами, обеспечивают вращение глазного яблока в глазнице.
Стенка глазного яблока состоит из трех оболочек, которые различаются по происхождению, строению и функциям (рис. 12.1, а).
- 1. Склера — наружная оболочка, образованная плотной соединительной тканыо. Она выполняет защитную функцию. Роговица — прозрачная передняя часть склеры, ее прозрачность обеспечивается упорядоченным расположением слоев соединительной ткани.
- 2. Сосудистая оболочка занимает среднее положение в стенке глазного яблока. В ней много кровеносных сосудов и пигментных клеток. Видимая часть сосудистой оболочки называется радужной оболочкой.
- 3. Сетчатка — наиболее внутренний слой стенки глазного яблока. В ней расположены фоторецепторы и система нейронов, обеспечивающих первичную обработку зрительных сигналов на периферическом уровне. Аксоны части нейронов (ганглиозных клеток) формируют зрительный нерв.
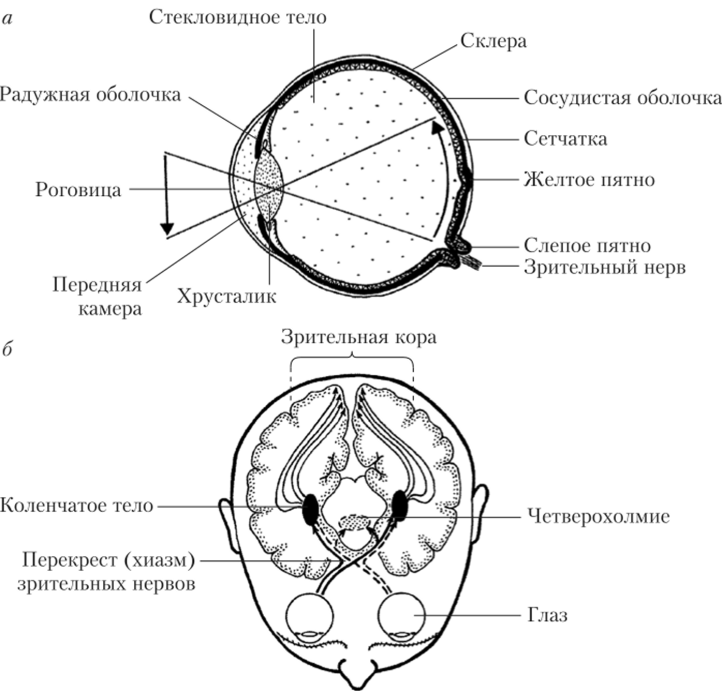
Рис. 12.1. Строение органа зрения (а) и основных зрительных нервных путей (б) В ходе эмбриогенеза глаза возникают как выпячивания боковых стенок промежуточного мозга. В результате образуются глазные пузырьки, сообщающиеся с мозгом с помощью глазных стебельков. Из последних формируются зрительные нервы, а из глазного пузырька — сетчатка. Между роговицей и радужной оболочкой располагается передняя камера глаза, которую заполняет прозрачная жидкость, свободно пропускающей свет к внутренней части глаза. В центре радужной оболочки находится отверстие, окруженное клетками гладкой мускулатуры, — зрачок (на рисунке не показан). Благодаря его способности менять свой диаметр регулируется количество света, попадающего на сетчатку.
Сразу за зрачком располагается прозрачная упругая линза — хрусталик, который состоит из клеток, потерявших ядра и органеллы и содержащих большое количество упорядоченно расположенного белков кристаллинов. Хрусталик не имеет кровеносных сосудов, в нем отсутствуют нервные волокна. Рост хрусталика прекращается в возрасте трех лет. В дальнейшем происходит постепенная потеря хрусталиком воды, в результате чего его объем несколько уменьшается. У пожилых людей упорядоченное расположение кристаллинов в клетках хрусталика нарушается, и хрусталик теряет прозрачность (катаракта).
Хрусталик подвешен на связках, которые крепятся к особой ресничной мышце, образованной гладкомышечными клетками. Эта мышца способна при сокращении изменять кривизну хрусталика, и его форма меняется от плоской к более выпуклой.
В результате изменения кривизны хрусталика возникает аккомодация — реакция, обеспечивающая четкое видение объектов, расположенных на разном удалении от глаз («наводка на резкость»). Рассматривание близких предметов требует более выпуклого хрусталика, что достигается увеличением напряжения ресничной мышцы. Если же нужно четко увидеть происходящее вдали — ресничная мышца расслабляется, связки натягивают хрусталик, и он становится плоским. Тонус ресничной мышцы регулируется парасимпатическим центром среднего мозга.
Стекловидное тело находится между хрусталиком и сетчаткой и представляет собой прозрачное содержимое глазного яблока.
Сетчатка по своему строению и функциям представляет собой нервный центр, в котором происходят первичная обработка зрительных сигналов и преобразование их в нервные импульсы, передаваемые в ГМ по аксонам зрительного нерва.
Сигналы в сетчатке передаются последовательно через цепочку из трех основных типов клеток: фоторецепторы (палочки и колбочки), биполярные клетки, ганглиозные клетки. Дополнительное взаимодействие между этими типами клеток обеспечивается горизонтальными и амакриновыми клетками (рис. 12.2, а).
Фоторецепторы — палочки и колбочки — составляют самый внутренний слой сетчатки. В следующем слое располагаются тела горизонтальных (образующих синапсы с фоторецепторами) и биполярных клеток.
Окончания аксонов биполярных клеток образуют сложные синапсы на дендригах ганглиозных клеток и отростках амакриновых клеток. Амакриновые клетки своими отростками образуют связи с другими амакриновыми клетками, а также с дендритами ганглиозных клеток, и иногда с дендритами биполярных клеток. Аксоны ганглиозных клеток образуют зрительный нерв, а их тела составляют внешний слой сетчатки.
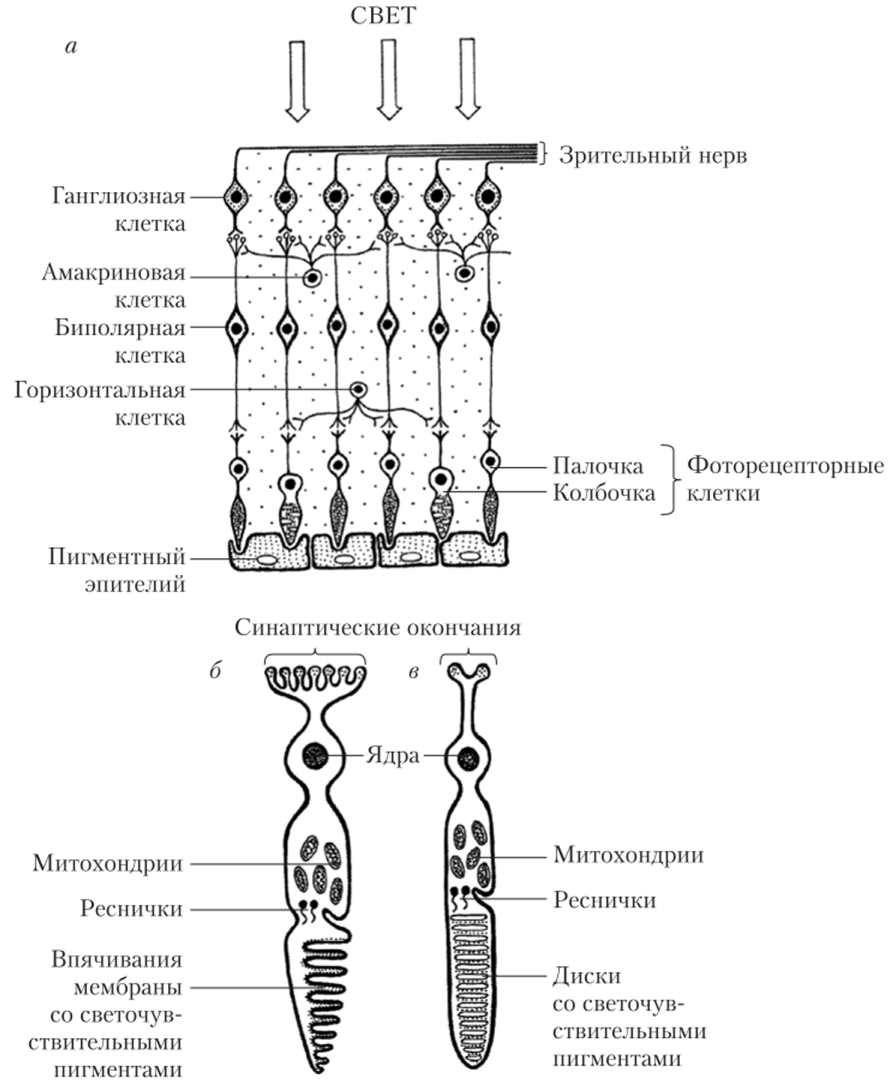
Рис. 12.2. Клеточное строение сетчатки (а), структура колбочки (б) и палочки (в) Фоторецепторная клетка (палочка или колбочка) состоит из ядерной области и примыкающих к ней с одной стороны наружным сегментом, в мембранах которого содержатся фотопигменты, а с другой — пресинаптическим окончанием, которое контактирует с отростками горизонтальных и биполярных клеток сетчатки (рис. 12.2, б> в). Зрительные пигменты обладают способностью менять свою форму, или конформацию (в конечном счете распадаясь), при попадании квантов света с определенной длиной волны, запуская каскад химических реакций в фоторецепторе, снижая секрецию медиатора из пресинаптического окончания, что в конечном счете, влияет на генерацию импульсов ганглиозными клетками сетчатки.
На первом этапе зрительного восприятия световая энергия трансформируется в изменение потенциала на мембране фоторецепторов. Этот процесс начинается в содержащих зрительный пигмент наружных сегментах палочек и колбочек. В палочках содержится пигмент родопсин, в колбочках — один из йодопсинов. У человека три типа йодопсинов: красно-, зеленои сине-чувствительный. Каждая конкретная колбочка содержит только один из йодопсинов, их совокупность обусловливает наличие цветового зрения. Родопсин реагирует на свет практически во всем видимом диапазоне, и поэтому реакция палочек на оранжевый свет не отличается от реакции на зеленый. При этом у палочек очень высокая светочувствительность. Такие рецепторы могут обеспечить только черно-белое зрение. Из-за этого в сумерках, когда света недостаточно для активации колбочек, палочки являются единственным источником зрительной информации. В условиях яркой освещенности палочки участвуют, прежде всего, в реакциях на движение и в четком выделении границ объектов.
Молекула зрительного пигмента состоит из белковой части — опсина (опсины различаются у палочек и разных типов колбочек), и хромофора — части, поглощающей свет. Хромофор одинаков у колбочек и палочек, по химическому составу он представляет собой альдегид витамина, А (ретиналь), способный принимать разные конформации. Недостаток витамина, А в пище приводит к ухудшению зрения в сумерках («куриная слепота»). Палочки и колбочки отличаются друг от друга по структуре и свойствам, однако основные принципы рецепции света у них одинаковы.
Наружные сегменты фоторецепторов содержат высокоэффективную молекулярную систему усиления светового сигнала. Электрофизиологическая регистрация сигналов от палочек и колбочек в темноте выявила входящий в наружный сегмент натриевый ток (темновой деполяризующий ток). Было показано, что в темноте фоторецепторы непрерывно секретируют медиатор (глутаминовую кислоту), действующий деполяризирующим или гиперполяризующим образом (в зависимости от типа рецепторов для медиатора) на постсинаптические мембраны отростков биполярных и горизонтальных клеток. Освещение фоторецепторной клетки приводит к закрытию каналов, обеспечивающих темновой ток. Возникающий гиперполяризующий рецепторный потенциал снижает секрецию медиатора в синапсе, вызывая гиперполяризацию или деполяризацию биполярных клеток.
В сетчатке человека около 140 млн палочек и 7 млн колбочек. Распределение палочек и колбочек по сетчатке неравномерно. Зона наибольшей остроты зрения расположена в самом центре сетчатки. Она содержит максимально плотно расположенные колбочки и носит название центральной ямки, или желтого пятна. На периферии сетчатки фоторецепторов значительно меньше, и это преимущественно палочки.
Распространенная патология, при которой наблюдаются нарушения цветового зрения, носит название дальтонизм. Это заболевание имеет генетическую природу и регистрируется у 7% мужчин и 0,5% женщин. Причиной дальтонизма является отсутствие одного из йодопсинов. По этой причине у дальтоника меняется восприятие цветов: он видит только два из трех основных цветов.
Видимое изображение фоторецепторы «описывают» как меняющуюся в пространстве и во времени совокупность красных, зеленых, синих и серых точек. Данная информация разделяется в результате дополнительной обработки клетками сетчатки на несколько параллельных потоков, которые в виде нервных импульсов поступают в ГМ по зрительному нерву. Работу сетчатки можно сравнить с работой матрицы фотоаппарата или сканера, которые преобразуют изображение в совокупность точек (пикселей) одинакового размера. При этом «пиксели» сетчатки имеют разный размер: самые мелкие — в области желтого пятна, а на периферии — значительно крупнее. Это способствует снижению объема информации, передаваемой в ЦНС, и экономит «вычислительный ресурс» мозга.
Зрительные центры головного мозга. Зрительный нерв образуют примерно 1 млн аксонов ганглиозных клеток сетчатки. После выхода из глазницы он направляется к зрительным центрам ГМ. Примерно 50% волокон зрительного нерва идет напрямую в «свою» половину мозга, а остальные образуют перекрест перед входом в промежуточный мозг. Этот нрекрест называется зрительной хиазмой (hiasma optica). После перекреста волокна зрительного нерва направляются к супрахиазменным ядрам гипоталамуса, верхним холмикам четверохолмия, нервным центрам задней части таламуса и первичной зрительной коре.
В гипоталамусе (в супрахиазменных ядрах) информация об интенсивности света и длительности освещенности используется для регуляции суточных ритмов организма.
В среднем мозге (в верхних холмиках четверохолмия) на основе поступающей от сетчатки зрительной информации осуществляется регуляция движений глаз (саккады), диаметра зрачка, формы хрусталика, а также обеспечивается запуск ориентировочного рефлекса в ответ на появление новых зрительных сигналов.
Нервные центры заднего таламуса (латеральные коленчатые тела) готовят зрительную информацию к поступлению в кору больших полушарий, контрастируя изображение с использованием контуров латерального торможения. Зрительная кора располагается в затылочной области больших полушарий и частично заходит на их медиальную поверхность (рис. 12.1, б). В зрительной коре происходит интеграция параллельных потоков зрительной информации и реализуется узнавание усложняющихся визуальных образов.
Первичная зрительная кора занимает самую заднюю затылочную область коры (поле 17). Чередующиеся группы нейронов реагируют на появление в иоле зрения по-разному ориентированных линий (по отношению к горизонту). Одна популяция нейронов отвечает максимальной активностью на горизонтальные отрезки, другая — на вертикальные, третья — на идущие под углом 45° и т. п.
Спереди от первичной находится вторичная зрительная кора. Она суммирует информацию о наличии нескольких отдельных линий, обеспечивая восприятие геометрических фигур. В этой области происходит объединение черно-белого потока (частично обусловленного палочковым сигналом) и потока сигналов о цвете (колбочковый сигнал). Тут сравнивается информация от правого и левого глаза (из-за наличия хиазмы такая информация попадает в одно полушарие и начинает обрабатываться еще в первичной зрительной коре), что позволяет оценить объем объектов и расстояние до них.
На границе затылочной, теменной и височной областей располагается третичная зрительная кора, синхронная работа целых групп в которой обеспечивает распознавание наиболее сложных и обобщенных признаков рассматриваемых объектов. Эта область коры у человека связана с узнаванием знакомых лиц, а также с восприятием написанного текста и чтением.
Повреждения первичной зрительной коры приводят к утрате отдельных участков в поле зрения и практически не компенсируются. Повреждения вторичной и третичной коры нарушают восприятие и узнавание зрительных образов, но хорошо компенсируются, поскольку свойства нейронов в этих областях являются результатом обучения.
Зрительная адаптация. Поскольку восприятие света приводит к распаду зрительных пигментов, то понятно, что интенсивное освещение вызывает в фоторецепторах быстрое снижение количества молекул фотопигментов. Такое «выцветание» фотопигментов компенсируется процессом повторного синтеза (ресинтеза) пигментов из продуктов их распада. Чем интенсивнее происходит распад пигментов, тем значительнее идет их ресинтез. В результате распад и ресинтез уравновешиваются на уровне, соответствующем степени освещенности. Так, на ярком солнечном свету содержание в палочках родопсина составляет не более 1—2% максимально возможного. При переходе в сумрак такого небольшого количества зрительного пигмента недостаточно для нормального зрительного восприятия. Человек при этом некоторое время ничего не видит, но в результате синтеза количество пигментов увеличивается, и через 10—30 мин наступает темновая адаптация — возможность различить предметы при очень низком уровне освещенности. Если снова переместиться в ярко освещенное место, пигменты в фоторецепторах начнут стремительно разлагаться (выцветать) и наступит временное перевозбуждение зрительной системы («ослепление»). В течение нескольких секунд количество фотопигмептов сильно снизится, произойдет световая адаптация и возможность видеть возвратится.
Темновой и световой адаптации способствуют рефлекторные изменения диаметра зрачка: количество света, поступающего на сетчатку, может изменяться почти в 20 раз. Ту же функцию обеспечивает и рефлекторное закрывание глаз при резком увеличении уровня освещенности.
Зрительные иллюзии. Фоторецепторы являются довольно «медлительными» сенсорными клетками, и реакция (гиперполяризующий рецепторный потенциал) на даже короткий световой стимул развивается 20—25 мс, а затем в течение такого же времени угасает. Это приводит к тому, что при повторах зрительных сигналов с частотой более 20 Гц они начинают сливаться, т. е. ответ на очередной кадр суммируется с ответами на предыдущий и последующий стимулы. В результате дискретный (мигающий) сигнал начинает восприниматься как непрерывный. Именно благодаря этому возможно создание «живых» кинои телеизображений, реально представляющих собой смену кадров с частотой 24 Гц и более.
Иллюзия цвета. Разнообразия цветов в реальности не существуют, а есть только сигналы от сине-, зеленои красно-чувствительных колбочек, реагирующих на электромагнитные волны разной длины. Однако наши внутренние «графические программы» представляют нашему сознанию информацию о том, сколько объект отражает (излучает) синих, зеленых и красных волн именно в виде «смешанных» светов: оранжевого, сиреневого, изумрудного и т. д.
Идея о том, что глаз видит всего несколько «чистых» цветов, а их слияние происходит уже в мозге, появилась еще в конце XVIII в. (эксперименты Томаса Юнга), а в середине XIX в. сыграла свою роль в возникновении импрессионизма, особенно такого его направления, как пуантель. Современные технические системы (цветные телевизоры и дисплеи) используют тот же принцип — создание сложного цвета за счет слияния трех «простых»: красного, зеленого, синего.
Иллюзия объема. Объем, конечно, существует реально. Но каждый наш глаз «сканирует» плоскую картинку. И далее вторичная зрительная кора сравнивает два плоских изображения и «вычисляет» объем. Эту систему легко обмануть — отсюда появление ЗЛ-картинок и ЗЛ-фильмов.
Иллюзии контекстного восприятия. В зависимости от окружения и мотивации мы опознаем разные образы на одном и том же изображении.
Движения глаз. Для успешного распознавания зрительных образов очень важны движения глаз. Они осуществляются при помощи трех пар черепных нервов (III, IV и VI), которые иннервируют шесть пар глазодвигательных мышц. Движения глаз управляются центрами, находящимися в верхних холмиках четверохолмия. Эти центры, в свою очередь, контролируются корой больших полушарий, мозжечком и базальными ганглиями.
Основные типы движений глаз — это саккады и плавные следящие движения.
Саккады — скачкообразные движения глаз длительностью 10—80 мс. Они сопровождают фиксацию взора на каком-либо объекте, слежение за объектом, рассматривание изображения. Амплитуда саккад может быть очень небольшой — всего несколько угловых минут, но может составлять более 90°. Длительность саккады не зависит от ее амплитуды, т. е. к моменту начала саккады мозг «просчитывает», насколько надо сдвинуть взор, и угловая скорость оказывается пропорциональна амплитуде.
Прослеживание движущегося объекта осуществляется благодаря плавным движениям глаз. Оно включается специальными нейронами — детекторами движения — в верхних холмиках четверохолмия. Минимальный порог скорости (абсолютный порог), запускающий слежение, примерно 5°/с, максимально возможная скорость слежения — 30—40°/с. Если эта скорость больше, возникают саккады. Начало слежения отстает от начала движения на 0,15—0,2 с, но скорость движения глаз сразу же соответствует скорости движения объекта. Следовательно, ЦНС осуществляет предварительное программирование движений глаз.
При рассматривании изображения происходит фиксация взора на наиболее информативных элементах — на контуре предмета, точках смены направления контура, при рассматривании лица (на глазах, губах и т. д.). Периоды фиксации сменяются саккадами. Иерархия выбора точек фиксации зависит от индивидуального опыта, мотивации, поставленной задачи.
При поражениях лобных долей, где находятся центры планирования движений, наблюдаются хаотичные фиксации взора. Анализ движений глаз, времени и последовательности фиксаций активно используется в современных нейромаркетологических исследованиях.