Контрольные вопросы.
Физиология растений в 2 т. Том 1

При незначительной плотности растений каждый лист получал бы больше света, и зависящая от освещенности интенсивность фотосинтеза и величина ЧПФ были бы выше, но при пересчете на площадь посева, плантации несомкнутый покров продуцирует меньше сухих веществ, чем сомкнутый. Однако если растения расположены слишком близко друг к другу и листья многократно перекрываются, то самым нижним из них… Читать ещё >
Контрольные вопросы. Физиология растений в 2 т. Том 1 (реферат, курсовая, диплом, контрольная)
- 1. Какие органы являются донорами? Какие органы называются акцепторами?
- 2. Какие вещества могут выходить из хлоропласта в цитозоль?
- 3. Какие вещества синтезируются в цитозоле из триозофосфатов?
- 4. Какие типы транспорта ассимилятов вы знаете?
- 5. Какой транспорт называется ближним и какой дальним?
- 6. По каким путям идет ближний транспорт? Какова его длина?
- 7. Как происходит загрузка флоэмиых окончаний?
- 8. Какое строение имеет ситовидная трубка?
- 9. Какую функцию выполняют сопровождающие клетки?
- 10. Что такое передаточные клетки? Какая у них функция?
- 11. Почему в ситовидных трубках должен быть замедлен обмен веществ (метаболизм)?
- 12. Какие вещества входят в состав флоэмного сока?
- 13. С какой скоростью идет дальний транспорт?
- 14. Какие внешние факторы влияют на скорость транспорта веществ по флоэме?
- 15. Когда отток ассимилятов происходит быстрее — днем или ночью?
- 16. Как объясняет транспорт веществ по флоэме теория массового тока под давлением?
- 17. Как объясняет транспорт веществ по флоэме электроосмотическая теория?
- 18. От чего зависит направление транспорта ассимилятов?
- 19. В каком направлении движутся вещества по флоэме?
- 20. Как влияют гормоны на скорость и направление транспорта ассимилятов по растению?
- 21. Как растение может регулировать скорость и направление оттока веществ?
- 22. Что такое каллоза? Как она влияет на транспорт ассимилятов?
- 23. Что влияет на распределение ассимилятов в растении?
Фотосинтез и урожай
Фотосинтез играет ведущую роль в формировании урожая. Говоря об урожае, важно различать два понятия: урожай биологический и урожай хозяйственный. Биологическим урожаем (УбиоЛ) называется масса органического вещества, образованного всеми растениями на 1 га почвы в течение вегетационного периода. Однако большая часть биологического урожая не используется человеком в качестве нищи или промышленного сырья. Хозяйственный урожай (Ухоз) — это доля сухого вещества, ради которого выращивают растения (плоды, семена, клубни и пр.):
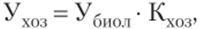
где Кхоз — коэффициент хозяйственной эффективности фотосинтеза, т. е. та часть сухого вещества, которая образует урожай.
Величина Кхоз зависит от культуры. Например, у зерновых она равняется 0,25—0,40 и даже 0,50; у сахарной свеклы — 0,5; у хлопчатника — 0,01. Кхоз может колебаться и у растений разных сортов одной культуры.
В среднем на планете биологический урожай равен 8 т/га, у сельскохозяйственных растений — 4 т/га. Однако его величина очень сильно отличается у разных растений: у С3-растений он может составлять 25 т сухого вещества в год с 1 га, у С4-растений — 55 т, а у сахарного тростника — 120 т.
Биологический урожай в умеренной зоне достигает 20—40 т/ra, в тропическом лесу — 100 т/га. Для человека важнее хозяйственный урожай.
Сухая масса растения на 90—95% состоит из органического вещества, источником которого является фотосинтез. Однако зависимость между фотосинтезом и урожаем очень сложна, изменчива и не всегда хорошо видна. Так, например, внесение минеральных удобрений увеличивает урожай в два, а в некоторых случаях — в три раза, хотя интенсивность фотосинтеза при этом не изменяется.
Величина биологического урожая определяется разностью между интенсивностью фотосинтеза и интенсивностью дыхания. На дыхание, без которого невозможна жизнь растения, в среднем расходуется до 15—25% ассимилятов. Однако дыхание нельзя рассматривать только как процесс расхода органического вещества. Во время дыхания образуется АТФ, необходимая для работы клеток, а следовательно, для создания урожая.
Первым, кто попытался связать с помощью математической формулы фотосинтез, дыхание и урожай, был известный русский физиолог Л. А. Иванов:
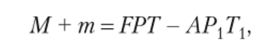
где М — конечный урожай, общая сухая масса, накопленная к моменту уборки; т — масса сухого вещества, синтезированного растением, но не вошедшая в конечный урожай; F — интенсивность фотосинтеза; Р — фотосинтезирующая поверхность, в основном поверхность листьев; Т — продолжительность работы фотосинтезирующего аппарата; FPT — общее количество образованного в процессе фотосинтеза органического вещества; А — интенсивность дыхания; Рх — масса всех дышащих органов; Тх — продолжительность жизни всех органов; АРХТХ — количество органического вещества, израсходованного на дыхание.
Для накопления сухого вещества интенсивность фотосинтеза должна быть в несколько раз выше дыхания. Фотосинтез идет только в зеленых клетках и только днем, а дыхание — во всех живых клетках днем и ночью. Поэтому величина Т всегда меньше величины Тх. Время Тх определяется временем жизни органов и всего растения.
Не все образовавшееся органическое вещество накапливается. Оно расходуется не только на дыхание, но и вымывается дождями, выделяется корнями, теряется при опаде органов (т). У некоторых растений, например у люцерны, до 25% ассимилятов тратится на образование цветков, завязей, плодов, которые при слабом освещении опадают раньше срока. При разреженных посевах опад уменьшается. В величину т входят и водорастворимые вещества: аминокислоты, сахара, органические кислоты, ионы, вымываемые из растений во время дождя, а особенно во время тропических ливней. Так, у сахарного тростника вымывается из листьев до 12% отложенных в них сахаров. Следовательно, значительная часть продуктов фотосинтеза не входит в урожай.
Для повышения урожая важно соотношение автотрофных (листья) и гетеротрофных (стебли, корни) органов. Растения, имеющие очень большую корневую систему по сравнению с надземной частью, тратят много веществ на дыхание, и поэтому побеги медленно растут. Деревья вначале ограничиваются образованием сравнительно малого количества листьев и строят прежде всего ветви и корневую систему. В дальнейшем развитии отношение надземной массы к корневой системе изменяется в пользу надземной части. Для сельскохозяйственных культур решающее значение имеет величина расхода ассимилятов на образование новых листьев или других длительно живущих частей растения.
Даже эта простая формула Иванова, показывает, что можно увеличить биологический урожай (М), применяя различные агротехнические приемы (полив, удобрения), используя высокоурожайные сорта, увеличивая интенсивность и продуктивность фотосинтеза.
Более детально связь урожая и фотосинтеза была разработана А. А. Ничиноровичем в 1955—1975 гг. (СССР) в его теории «Фотосинтетическая продуктивность растений». По этой теории, биологический урожай (Убиол) равен сумме суточных приростов сухого вещества в течение всего вегетационного периода:
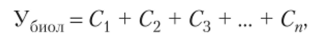
где С — прирост сухой массы, кг/га; п — число суток.
Прирост сухой массы (С) можно определить по формуле.
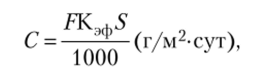
где F — интенсивность фотосинтеза, мг С02/м2ч; S — площадь листьев; Кэф — коэффициент, позволяющий перейти от количества поглощенного С02 к величине накопленного сухого вещества; он равен 0,64, т. е. 1 кг поглощенного углекислого газа соответствует 0,64 г углеводов.
Как уже говорилось, количество граммов сухого вещества, накопленного одним квадратным метром листовой площади растения за сутки, характеризует чистую продуктивность фотосинтеза (ЧИФ). Она складывается из количества граммов сухого вещества, образованного во время фотосинтеза, минус количество граммов сухого вещества, разрушенного при дыхании. Чем больше эта разница между интенсивностью фотосинтеза и интенсивностью дыхания, тем больше чистая продуктивность фотосинтеза. Если интенсивность дыхания высока, то ЧПФ может быть низкой и при большой интенсивности фотосинтеза. Чистая продуктивность фотосинтеза сильно отличается у растений с разным типом фотосинтеза. Так, у С4-растений она равна 4—5 г/дм2-сут, у С3-растений — до 2, а у САМрастений — всего 0,015—0,018.
По теории фотосинтетической продуктивности растений, главными факторами, определяющими величину биологического урожая, являются:
- 1) размер площади фотосинтетического аппарата, главным образом листьев;
- 2) скорость его формирования;
- 3) интенсивность и продолжительность его работы;
4) соотношение между процессами образования и расходования органического вещества.
Прямая связь между урожаем и интенсивностью фотосинтеза наблюдается не всегда. Как уже было сказано (см. параграф 4.6), наибольшая интенсивность фотосинтеза — у растений пустынь и полупустынь, а растения растут там крайне медленно. В тропиках условия для роста благоприятны, но интенсивность фотосинтеза низкая. Средняя скорость фотосинтеза в полевых условиях раз в пять меньше потенциальной, а в экстремальных условиях нередко падает до нуля. Путь к повышению средней устойчивой интенсивности фотосинтеза — создание оптимальных условий для растений. Это задача агротехники.
Большое значение имеют скорость формирования и конечная площадь листьев. Если у растений формируется маленькая листовая поверхность, то урожай низкий; если много листьев, то нижним листьям может не хватать света, и интенсивность фотосинтеза будет на уровне компенсации.
Для характеристики площади листьев обычно используется величина листового индекса. Индекс листовой поверхности (ИЛИ) — это отношение суммарной поверхности всех листьев к площади почвы, занимаемой данными растениями. Так, если ИЛП равен 3, значит поверхность всех листьев в три раза больше площади почвы, на которой растут растения. Оптимальная для посева площадь листьев зависит от их расположения на стебле. Например, высокоурожайные сорта сахарной свеклы имеют приподнятые над землей розетки листьев, а низкоурожайные — распластанные по земле (см. рис. 4.39). Чем вертикальнее расположены листья, тем меньше верхние листья затеняют нижние и гем больше может быть индекс листовой поверхности. У растений, имеющих прилегающие к земле розетки листьев, максимальная интенсивность фотосинтеза в посеве достигается уже при ИЛП, равном 1,0—1,5. В то же время растения с узкими листьями, расположенными на побеге более или менее вертикально и равномерно, имеют высокую интенсивность фотосинтеза при большем значении ИЛП.
В естественных условиях ИЛП варьирует в зависимости от плотности насаждения между величинами 1 для молодых саженцев и до 8—9 для густых хвойных насаждений. В летне-зеленых смешанных насаждениях лиственных пород в Японии и в вечнозеленых галерейных лесах Таиланда он достигает больших величии: 13,7 и 16,6. Для сельскохозяйственных культур ИЛП тоже сильно варьирует и в большой степени зависит от агротехники. Так, у клевера оптимальная величина ИЛП равна 3—4, у пшеницы — до 7. У тропических злаков индекс листовой поверхности теоретически может достигать 15, так как листья расположены вертикально в течение большей части жизненного цикла, что позволяет лучше поглощать свет даже при формировании большой листовой поверхности. Практически же ИЛП обычно колеблется от 2,0 до 3,7.
В условиях плохой агротехники хозяйственный урожай лимитируется невысокими значениями ИЛП (1—1,5). В этом случае повышение урожая зависит от увеличения площади листьев, ИЛП должен равняться 3—4.
Однако увеличение площади листьев благоприятно для урожая лишь до определенного предела. В среднем для умеренной зоны оптимальной считается 5 га листьев на 1 га пашни, для влажных тропиков — 10 га листьев. При дальнейшем ее увеличении начинается взаимное затенение листьев, ухудшается поступление света к нижним листьям, что приводит к уменьшению чистой продуктивности фотосинтеза. Слишком большое увеличение листовой поверхности в сельскохозяйственном посеве или посадке может привести к уменьшению Кхоз, так как листья будут затенять друг друга, да и чем больше листьев, тем больше ассимилятов тратится на их образование и дыхание. Индекс листовой поверхности — один из важных показателей, определяющих количество света, поглощаемого растениями данной популяции.
Биологическая продуктивность посева зависит от его фотосиитетического потенциала (ФПП) — суммы величин площади листьев в м2/га за каждые сутки в течение всего вегетационного периода. Его величина варьирует у разных растений от 0,5 до 5 млн м2/га*сут.
Большое значение имеет густота стояния растений. Увеличение количества растений может привести к снижению урожая, из-за того что растения станут затенять друг друга. Для каждой культуры и для каждого района ее возделывания нужно найти оптимальную величину фотосинтезирующей поверхности.
В посевах, на плантациях происходит саморегуляция величины листовой поверхности за счет самоизреживания — гибели части листьев или части растений. В результате можно посеять много семян, а общая площадь листьев всех растений будет такая же, как и при меньшей норме высева. При большой облиственное™ побегов у нижних листьев интенсивность фотосинтеза из-за затенения может быть ниже интенсивности дыхания. В результате эти листья не только не участвуют в создании урожая, но еще получают ассимиляты от других листьев для дыхания и построения своих клеток. Формирование оптимальной листовой поверхности в посеве, на плантации — основной метод управления урожаем.
Для создания высокого урожая важна не только величина листовой поверхности, но и скорость ее формирования, т. е. как быстро в течение вегетации она достигнет своей максимальной величины, а также сколько времени каждый лист будет находиться в физиологически активном состоянии и какова продолжительность жизни листа. Например, у одних сортов сахарного тростника площадь листьев достигает максимальной величины к 4-месячному возрасту, затем снижается и до уборки стеблей держится на постоянной величине, так как образование новых листьев компенсируется количеством засохших; у других сортов листовой индекс достигает максимальной величины к 10-месячному возрасту и сохраняется на этом уровне до рубки. Скорость формирования листьев зависит от вида растения, их снабжения водой, минеральными веществами, от освещения и температуры. Нужно добиваться быстрого развития листовой поверхности в начале вегетационного периода, но следует помнить, что чрезмерное число листьев нежелательно, они затеняют друг друга.
На поступление света в посев влияют высота стояния солнца, погода, количество растений в поле, сомкнутость посева, облиственность каждого растения, расположение листьев на побеге.
На своем пути через растительный покров свет должен пройти сквозь последовательные слои листьев. Больше солнечного света поглощают первые два листа. В общей сложности шесть самых верхних листьев перехватывают 70% света, падающего на поверхность посева или посадки (табл. 4.8). Следовательно, в основном эти листья образуют органическое вещество побега или растения. Так как число листьев на растении с возрастом изменяется (образуются новые листья, но отмирают нижние), то распределение света в посеве на протяжении вегетационного периода тоже изменяется.
Таблица 4.8
Количество света, падающего на отдельные листья сахарного тростника, кал/см2*мин.
Номер листа от верхушки побега | Количество света | Номер листа от верхушки побега | Количество света |
0,61 | 0,39 | ||
0,62 | 0,32 | ||
0,57 | 0,17 | ||
0,49 | 0,14 | ||
0,46 | 0,13 |
При незначительной плотности растений каждый лист получал бы больше света, и зависящая от освещенности интенсивность фотосинтеза и величина ЧПФ были бы выше, но при пересчете на площадь посева, плантации несомкнутый покров продуцирует меньше сухих веществ, чем сомкнутый. Однако если растения расположены слишком близко друг к другу и листья многократно перекрываются, то самым нижним из них не хватает света, чтобы поддерживать фотосинтез выше точки компенсации. В густых посадках нижние листья, а иногда и средние, из-за самозатенения совсем не фотосинтезируют и тратят на дыхание ассимиляты других листьев. В этом случае биологический урожай с единицы площади постепенно снижается. Одновременно надо иметь в виду, что увеличение площади листьев приводит к увеличению транспирирующей поверхности растений, поэтому при недостаточном водоснабжении образование новых листьев тормозится. Отсюда ясно, что повышение продуктивности за счет увеличения листового индекса имеет определенный предел.
Продолжительность работы фотосинтезирующего аппарата складывается из продолжительности светлого периода суток, жизни самого растения и отдельного листа. Более урожайными являются сорта, у которых листья рано формируются и долго живут. Высокогорные растения малопродуктивны, так как у них интенсивность фотосинтеза выше компенсационной точки только утром, т. е. листья работают с большой интенсивностью недолго. В умеренном климате позднеспелые сорта урожайнее, так как у них фотосинтезирующий аппарат дольше работает. Продолжительность работы листьев у скороспелых сортов значительно меньше, чем у позднеспелых. Поэтому в районах, где вегетационный период не прерывается неблагоприятными условиями (засуха, пониженные температуры), больший урожай дают позднеспелые сорта. Озимые культуры зерновых более урожайны, чем яровые (см. параграф 7.3), так как их ассимиляционный аппарат работает дольше в оптимальных условиях.
Как увеличить продолжительность работы листьев? Нужно увеличить продолжительность жизни хлоропластов. Для этого необходимо создать для растений оптимальные водный режим и азотное питание. Время жизни тилакоидов также можно увеличить, обработав растения фитогормоном (кинетином) или синим светом.
Растения могут накопить достаточное количество сухого вещества даже при умеренной интенсивности света, если благоприятный для фотосинтеза период достаточно продолжителен.
Итак, наибольшие урожаи могут быть получены при быстром развитии оптимальной площади листьев, увеличении продолжительности их работы в течение суток и всего вегетационного периода, максимальном фотосинтезе и больших суточных приростах органического вещества. Все эти показатели варьируют в зависимости от культуры, сорта и условий выращивания. Всего этого можно достичь с помощью агротехники и селекции, нацеленной на повышение интенсивности фотосинтеза, скорости и направления оттока ассимилятов.
Биологический и хозяйственный урожаи тесно связаны. Величина хозяйственного урожая всех агрофитоценозов равна 700 млрд т в год. Этого недостаточно для удовлетворения всех потребностей человечества.
Для получения наибольшего хозяйственного урожая важно повысить величину коэффициента хозяйственного использования (Кхш). Это можно сделать, направляя отток ассимилятов в те органы, которые составляют урожай. Для чего нужно умело использовать различные агротехнические приемы, а также регуляторы роста. Для озимой пшеницы установлено, что урожайные сорта отличаются более высоким содержанием некоторых гормонов.
Селекционеры должны вывести новые сорта с высокими значениями коэффициента хозяйственного использования (Кхоз). Например, карликовые формы плодовых деревьев, злаков; сорта, быстро формирующие оптимальную листовую поверхность, с быстрым оттоком ассимилятов в запасающие органы. Создание высокопродуктивных короткостебелыibix (карликовых) форм позволяет растениям направлять большую часть ассимилятов на формирование хозяйственно ценных органов. При этом значительно возрастает коэффициент хозяйственного использования. Уже получены низкорослые сорта риса, пшеницы, которые не полегают, что также повышает их ценность.
У злаков значительная часть ассимилятов, используемых для формирования зерновки, образуется в стебле, в чешуях соцветия, остях, что особенно характерно для ячменя и пшеницы. У европейских пшениц доля участия фотосинтеза колоса в образовании зерновок меньше, чем у австралийских и индийских пшениц. Правда, для формирования хорошего колоса с большим количеством колосков и цветков нужны органические вещества, а они поступают из листьев. Следовательно, необходимо разумное соотношение органов.
Еще один путь повышения урожайности — увеличение процента использования фотосиптетически активной радиации. В естественных условиях растения используют 0,5—2% поглощенной энергии на фотосинтез, в искусственных системах — до 10%. КПД использования ФАР можно повысить с помощью:
- — создания агроценозов с максимально возможным ассимиляционным потенциалом (произведение площади листьев в посеве на длительность их функционирования);
- — формирования определенной пространственной организации агроценоза (общий габитус растения, размеры и формы листьев, их ориентация), что обеспечивает более равномерное распределение света в агроценозе и снабжение энергией всех ярусов листьев;
- — обеспечения растений С02, водой, элементами минерального питания.
Эволюция в природе и селекция шли не по пути изменения самого фото;
сиитетического аппарата, а по пути перераспределения продуктов фотосинтеза внутри самого растения: менялась скорость оттока ассимилятов, соотношение автои гетеротрофных органов, способность к повторному использованию органических веществ (реутилизация). Эволюция шла не на клеточном, а на органном и организменном уровнях, поэтому в ходе эволюции клетка почти не менялась. Теперь селекционеры должны обратить внимание на структуру и работу хлоронластов, т. е. перейти на клеточный и субклеточный уровни изменения фотосинтезирующего аппарата.