Аккумуляция тяжелых металлов орнитофауной

По данным дисперсионного анализа именно содержание Pb, Ni, Cd, Мо, Hg, Сг достоверно различается у птиц различного вида. Эти различия прямо связаны со структурой пищевых рационов изученных видов. Известно, что летние и осенние корма тетеревиных сходны. Наибольшие отличия характерны для зимнего периода, когда глухарь переходит на питание хвоей сосны, тетерев — концевыми побегами, почками… Читать ещё >
Аккумуляция тяжелых металлов орнитофауной (реферат, курсовая, диплом, контрольная)
Использование птиц в мониторинге состояния природной среды имеет давние традиции. Способность накапливать тяжелые металлы в организме дала основание использовать птиц и их органы в качестве биоиндикаторов загрязнения среды металлами.
Химические свойства элементов определяют форму их аккумуляции в организме, особенности метаболизма и органы преимущественного накопления. Многие элементы (Си, Cd, Pb, Hg, Со, Mn, As) концентрируются в органах выведения: печени и почках. Ряд остеотропных металлов (Pb, Zn, Al, Mn) накапливается в костной ткани. Органические соединения ртути (метили диметилртуть) депонируются в жировой ткани, мозге и в мышцах. В перьевом покрове накапливаются Zn, Сг, Си, Ni, Hg (Добровольская, 1987; Лебедева, 1999; и др.).
Содержание тяжелых металлов в организме птиц, обитающих на загрязненных территориях, коррелирует с их концентрацией во внешней среде. К таким видам относятся мухоловка-пеструшка, синица рода Parus, полевые воробьи, мохноногий сыч, серая куропатка и другие. Концентрация свинца в скелете взрослых диких птиц, превышающая 5 мкг/г, свидетельствует о загрязненности этим металлом среды их обитания.
Закономерности накопления тяжелых металлов в организме зависят от их химических свойств и физиологической роли. Выявлены положительные корреляции между концентрациями свинца, кадмия, ртути, мышьяка, а также в ряде случаев — меди в окружающей среде и в организме. В то же время тесные взаимосвязи между концентрациями цинка, железа в организмах птиц и окружающей среде отсутствуют, что, вероятно, связано с гомеостатическими механизмами. В качестве примера приведем уровни подвижных форм некоторых металлов в почвах и их концентрации в тканях птенцов (табл. 5.9).
Таблица 5.9
Концентрации металлов в почве (подвижные формы) и печени птенцов мухоловки-пеструшки в разных зонах загрязнения (мкг/г воздушно-сухой массы) (по: Бельский и др., 2005).
Субстрат. | Металл. | Зона токсической нагрузки. | ||
фоновая. | буферная. | импактная. | ||
Почва. | РЬ. | 67,3 ± 4,9. | 283,2 ± 14,1**. | 639,1 ± 43,2**. |
Cd. | 2,0 ± 0,2. | о. 1+. о со.
| 6,5 ± 0,4**. | |
Си. | 86,9 ± 7,6. | 862,4 ± 63,9**. | 3769,6 ± 151,9**. | |
Zn. | 135,2 ± 18,3. | 366,0 ± 34,2**. | 241,8 ± 11,8**. | |
п | ||||
Печень птенцов. | РЬ. | 5,5 ± 0,9. | 10,1 ± 1,7*. | 21,7 ± 2,8**. |
Cd. | 1,0 ± 0,1. | 1,0 ± 0,1. | 2,6 ± 0,3**. | |
Си. | 16,1 ± 1,0. | 14,9 ± 1,3. | 20,2 ± 2,2. | |
Zn. | 92,5 ± 6,2. | 125,8 ± 11,0*. | 145,3 ± 8,8**. | |
п | ||||
- * Отличия от фонового показателя значимы: р < 0,05.
- ** — р < 0,001.
Содержание металлов в печени птенцов мухоловки-пеструшки изменяется в градиенте токсической нагрузки аналогично их уровням в почве, однако кратность изменения этих показателей в градиенте загрязнения у птиц меньше, чем в почве: свинца в 3,9 раза, кадмия в 2,6 раза, меди и цинка в 1,3 и 1,6 раза соответственно. Это связано со снижением концентраций ряда металлов в процессе их миграции по пищевым цепям, а также с наличием в организме животных механизмов регуляции физиологически значимых элементов (среди названных металлов — медь и цинк). Сходные различия в накоплении в печени птенцов мухоловки-пеструшки мышьяка, кадмия, ртути и свинца — с одной стороны, и меди и цинка — с другой, отмечены в окрестностях металлургического завода в Швеции (Nyholm, 1995).
Видовая специфика накопления тяжелых металлов в тканях и органах птиц определяется структурой рациона, положением вида в трофической цепи, способом и местом кормодобывания, особенностями метаболизма. Так, концентрация Cd у растительноядных птиц на 1—2 порядка выше, чем у хищных и всеядных. Из растительноядных наиболее высокие концентрации Cd обнаружены у глухаря (почки содержат 43 мкг/г, печень 4,4 мкг/г сырого вещества) и лебедя-шипуна (в почках 42 мкг/г, в печени 12 мкг/г). Концентрации мышьяка в организмах хищных и рыбоядных птиц больше по сравнению с фитофагами, полифагами и энтомофагами. Накопление свинца в скелете интенсивнее протекает у мелких видов по сравнению с крупными, у наземных по сравнению с водными (Лебедева, 1999).
В качестве примера приведем анализ костных тканей птиц двух семейств: тетеревиных (Tetraonidae, Galliformes) и совиных (Strigidae, Strigiform.es). В сборах представлены следующие виды: глухарь (Tetrao urogallus L.), рябчик (Tetrastes bonasia L.), тетерев (Lyrurus tetrix L.) (Tetraonidae), а также неясыти: бородатая {Strix nebulosa Forster) и длиннохвостая {Strix uralensis Pall.) (Strigidae). Перечисленные виды оседлы, и, следовательно, химический состав их организма отражает уровни элементов в объектах среды конкретной локальной территории. Выбор двух семейств связан с тем, что они представляют разные трофические уровни: консументы первого (фитофаги) и второго (хищники) порядков. Следует подчеркнуть, что различия химического состава у этих двух групп обусловлены в первую очередь их положением в трофической цепи.
Проведен элементный анализ содержания 10 металлов в скелете тетеревиных и сов. Пробы птиц собраны в 13 точках Урала: от Приполярного до Южного. Широкий разброс точек сбора проб дал возможность оценить географическую специфику локальных популяций изучаемых видов. Большая часть птиц была отстреляна охотниками в осенний период, а часть — обнаружена в виде костно-перьевых остатков. В анализе использовали главным образом цевки птиц, реже — кости таза.
Мера дифференциации видовых выборок определялась при дискриминантном анализе с использованием квадратичных дистанций Махаланобиса. При анализе особенностей накопления элементов птицами разных трофических уровней рассчитывались средние значения элементов в двух выборках: в одну были объединены три вида тетеревиных, а в другую — оба вида сов. Сравнение средних концентраций элементов в скелете у двух групп консументов проводилось попарно с использованием t-критерия Стьюдента.
Пространство 10 показателей, соответствующих концентрациям химических элементов, отображается на плоскости двух компонент, на которой исходные данные представлены в виде соответствующих точек. Расстояние между ними характеризует близость изучаемого минерального состава костных тканей всех видовых групп птиц.
Полученные результаты позволили объединить неразличающиеся виды и продолжить дискриминантный анализ на трех выборках: глухарь и тетерев, рябчик и оба вида сов. Анализ объединенных выборок показал, что выделенные нами группы видов удовлетворительно дифференцируются по показателю квадратичных дистанций Махаланобиса при уровне значимости р < 0,05.
На рис. 5.24 представлено расположение объединенных выборок микроэлементного состава костных тканей на плоскости дискриминантных функций (95% доверительные эллипсоиды). Качество выполненной дифференциации можно охарактеризовать эффективностью классификации исходных выборок концентраций металлов, выполненной в рамках дискриминантного анализа и составляющей для совместной выборки глухаря и тетерева 92,3%, для выборки проб рябчика — 70,0%, для обоих видов сов — 84,2%. Общая вероятность правильного разделения таких выборок составляет 85,4%.
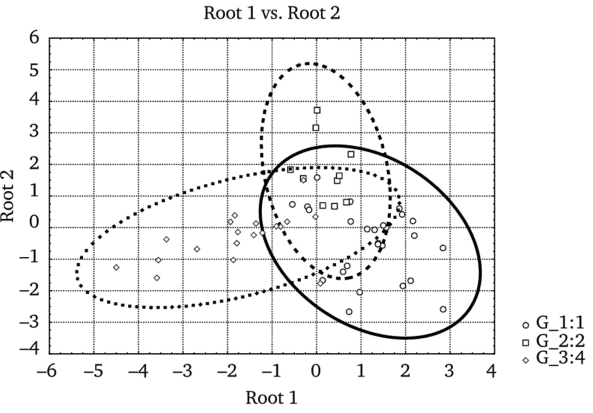
Рис. 5.24. Расположение объединенных выборок микроэлементного состава костных тканей птиц на плоскости дискриминантных функций (95% доверительные эллипсоиды):
G_l:l — объединенная выборка глухарей и тетеревов; G2:2 — выборка рябчиков; G3:4 — объединенная выборка сов Проведенный анализ позволил объединить данные с учетом особенностей накопления элементов птицами разных трофических уровней. С одной стороны, это тетеревиные (консументы I порядка), с другой — совы (консументы II порядка). Попарное сравнение средних концентраций элементов в скелете у двух трофических групп показало, что уровни Mn, Fe, Си и Zn статистически не различаются. Накопление двух металлов в костной ткани у птиц-фитофагов было больше, чем у хищных птиц. Так, средняя концентрация Ni составила соответственно.
1,44 ± 0,44 и 0,50 ± 0,25 мкг/г воздушно-сухой массы, Cd — 0,13 ± 0,05 и 0,006 ± 0,002 мкг/г. У сов статистически значимо в большей степени, чем у тетеревиных, накапливались РЬ и Мо. Среднее содержание РЬ составило 0,92 ± 0,49 мкг/г у тетеревиных, 3,93 ± 1,07 мкг/г у сов, а Мо соответственно 0,02 ± 0,02 и 2,27 ± 1,00 мкг/г. Различия в накоплении Сг находятся на грани статистической значимости (р < 0,1): 2,55 ± 0,52 и 3,27 ± 0,35 мкг/г у консументов I и II порядков соответственно.
Между видами не замечено значимых различий концентраций Мп, Fe, Си и Zn в костных тканях. Как указывалось выше, эти элементы выполняют важнейшие физиологические функции, и следует предполагать, что механизмы минерального гомеостаза контролируют постоянство их содержания в тканях и органах птиц. По этой причине перечисленные микроэлементы не обладают статистически значимой видовой спецификой. Видовые различия в накоплении следует ожидать в отношении остальных элементов, не относящихся к физиологически необходимым и по этой причине способных к пассивному транспорту через стенку ЖКТ. Поэтому уровни таких элементов в костных тканях птиц отражают их содержание в пищевых рационах.
По данным дисперсионного анализа именно содержание Pb, Ni, Cd, Мо, Hg, Сг достоверно различается у птиц различного вида. Эти различия прямо связаны со структурой пищевых рационов изученных видов. Известно, что летние и осенние корма тетеревиных сходны. Наибольшие отличия характерны для зимнего периода, когда глухарь переходит на питание хвоей сосны, тетерев — концевыми побегами, почками и сережками берез, а рябчик — на аналогичный веточный корм, включающий широкий набор лиственных деревьев и кустарников. При всех различиях зимнего рациона тетерева и глухаря у них имеется общее — использование в пищу тетеревами молодых шишечек и хвои сосны, характерное для северных районов Евразии. Рябчик, в наибольшей степени отличающийся своим рационом от этих двух видов, обладает и наименьшим сходством с ними по химическому составу. Таким образом, наблюдаемые различия в химическом составе разных видов тетеревиных связаны с особенностями их питания. Сходные пищевые рационы сов обусловливают единство химического состава тканей рассмотренных нами двух видов этих птиц.
Описываемое явление в природных условиях усложняется не только комплексом экологических факторов, формирующих состав пищевых рационов, но и химической формой элементов, поступающих в ЖКТ. Именно это обстоятельство в конечном счете определяет эффект увеличения либо уменьшения концентрации конкретного химического элемента при переходе к следующему трофическому уровню.
В нашем случае лишь содержание РЬ и Мо в костной ткани сов достоверно превышало таковое у тетеревиных. Обнаруженное уменьшение уровней Cd у хищных птиц по сравнению с фитофагами согласуется с данными ряда авторов о преимущественном накоплении этого элемента у растительноядных животных.
Различия между трофическими уровнями в концентрировании разных элементов также неодинаковы. При переходе на следующий трофический уровень содержание Ni и РЬ меняется в 3—4 раза, a Cd и Мо — в 20 и более раз. Средние концентрации других элементов варьировали в значительно меньшей степени. Таким образом, поведение конкретных элементов в биогенном круговороте строго специфично.
Несмотря на очевидное значение состава пищевых рационов в формировании токсической нагрузки, имеются данные о роли в этом процессе систематического положения птиц. В оперении птиц выделено две группы микроэлементов: сопряженные (Си, Zn, Mn, Ni и Со) и несопряженные (Cr, V, Ti, Mo, Fe, Ag). Для каждого таксона автор выделяет характерную группу элементов, содержание которых в перьях отражает систематическое положение птиц (Добровольская, 2002).
Другие исследования показали, что если уровни марганца, цинка, меди, железа в костях птиц Эфиопии не связаны с видовой принадлежностью, то накопление алюминия в тех же тканях видоспецифично (Лебедева, 1999).
Физиологические особенности обусловливают половую специфику аккумуляции тяжелых металлов в организме и чувствительности к их токсическому действию. Накопление свинца выше у самок сизого голубя и американской пустельги по сравнению с самцами. Это связано с более выраженной аккумуляцией Са и других остеотропных элементов в организме самок перед яйцекладкой. Для неостеотропных элементов половая специфика накопления либо не выражена (вяхирь, кваква, чайка-хохотунья, утиные), либо противоположна у разных видов. Так, у самок американской черной утки концентрация кадмия в печени ниже, чем у самцов, а у кряквы и морской чернети — наоборот. В перьях воротничкового рябчика (Bonasa umbellus) не выявлены зависимости накопления Си, Ni и Fe от пола и возраста птиц.
Особенности метаболизма определяют и возрастную специфику накопления тяжелых металлов в организме птиц. В период интенсивного роста тела и минерализации скелета происходит быстрое накопление металлов в организме молодых птиц. С возрастом поступление и выведение элементов из организма приходят к равновесию. Таким образом, в целом исходно низкие концентрации тяжелых металлов, присущие организму птенцов, с возрастом увеличиваются. Наиболее ярко это выражено для остеотропных элементов, депо которых — скелет — окончательно формируется и насыщается минеральными веществами лишь у взрослых особей. Увеличение с возрастом концентраций РЬ и Cd в перьях отмечено у сорок, но в оперении молодых выше уровни Zn, Си, Fe. Результаты исследований в Норвегии показали, что Cd в печени и почках молодых белых куропаток быстро накапливается в течение первой зимы, а затем, после достижения порогового уровня, его аккумуляция прекращается.
Концентрации тяжелых металлов в яйцах птиц коррелируют с их содержанием в организмах самок. Однако количество металлов, поступающих в яйца из организма самки, значительно ниже их аккумуляции в печени и почках. При увеличении содержания свинца в 5,6 раза в рационе птиц импактной зоны по сравнению с фоновой концентрация этого элемента в скорлупе изменяется лишь в 1,2 раза. Для Си подобные показатели составляли 6,5 и 2 раза. Очевидно, существуют физиологические механизмы, ограничивающие поступление тяжелых металлов в репродуктивные органы.