Взаимодействие гормонов с клетками-мишенями

Ядерные рецепторы, регулирующие синтез белка, характерны, например, для половых гормонов (андрогены, эстрогены), гормонов щитовидной железы (тиреоиды), глюкокортикоидов и минералокортикоидов надпочечников. Эти гормоны легко проникают через плазматическую мембрану в клетку, где в цитозоле связываются с рецепторным белком и транспортируются в ядро. В неактивном состоянии подсемейство… Читать ещё >
Взаимодействие гормонов с клетками-мишенями (реферат, курсовая, диплом, контрольная)
Взаимодействие гормона с клеткой-мишенью начинается с того, что гормон должен сформировать гормон-рецепторный комплекс, т. е. связаться с рецептором этой клетки. Клетки, лишенные рецепторов к какому-либо гормону, не способны реагировать на гормональное воздействие. Взаимодействие гормона и рецептора инициирует начало каскада ферментативных реакций, каждая стадия которого становится все более мощной, поэтому даже малая концентрация гормона способна давать существенный эффект.
Рецепторы гормонов являются крупными белками, и каждая чувствительная к гормону клетка может иметь от 2000 до 100 000 таких рецепторов. Рецепторы высокоспецифичны для конкретного гормона, что предопределяет тип гормона, который сможет оказать влияние на определенную ткань.
Подавляющее число рецепторов к гормонам локализуется следующим образом:
- — рецепторы к липофильным гормонам, хорошо проходящим в клетку (стероидные и тиреоидные гормоны, гормональные формы витаминов, А и D), являются ядерными и непосредственно участвуют в регуляции экспрессии определенных генов;
- — рецепторы к соединениям, не проникающим через клеточную мембрану (белково-пептидные гормоны, простагландины, катехоламины), располагаются на клеточной мембране. В результате их взаимодействия с гормоном запускаются внутриклеточные сигнальные каскады, что приводит к изменению степени фосфорилирования эффекторных белков или транскрипционных факторов.
Мембранные рецепторы подразделяют на обладающие собственной ферментативной активностью (рецепторные тирозинкиназы и гуанилатциклазы) и на не обладающие собственной ферментативной активностью (сопряженные с G-белками).
Последовательность событий, необходимых для развития гормонального эффекта, представлена на рис. 1.9.
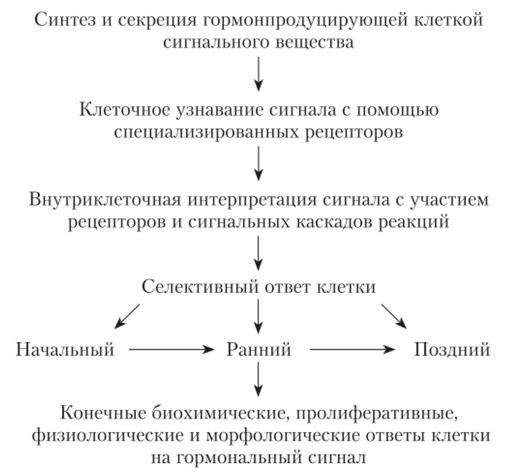
Рис. 1.9. Этапы развития гормональных эффектов.
Ядерные рецепторы, регулирующие синтез белка, характерны, например, для половых гормонов (андрогены, эстрогены), гормонов щитовидной железы (тиреоиды), глюкокортикоидов и минералокортикоидов надпочечников. Эти гормоны легко проникают через плазматическую мембрану в клетку, где в цитозоле связываются с рецепторным белком и транспортируются в ядро. В неактивном состоянии подсемейство глюкокортикоидных, протестероновых, эстрогеновых и андрогеновых рецепторов бывает связано с белком теплового шока (англ, heat-shock protein) HSP-90.
При связывании гормона с рецептором белок теплового шока отделяется от последнего, комплекс белок — рецептор связывается с ДНК в специфических участках, называемых элементами гормонального ответа, которые располагаются по направлению транскрипции за сайтами начала транскрипции. В результате транскрипция и синтез белка нарушаются. Рецепторы тиреоидных гормонов и ретиноевой кислоты в неактивном состоянии не связаны с белком теплового шока и могут присоединяться к своим элементам гормонального ответа на молекуле ДНК в отсутствие гормонов, подавляя транскрипцию. Активация рецепторов, реализующих действие гормонов, по-видимому, происходит в результате фосфорилирования (рис. 1.10).
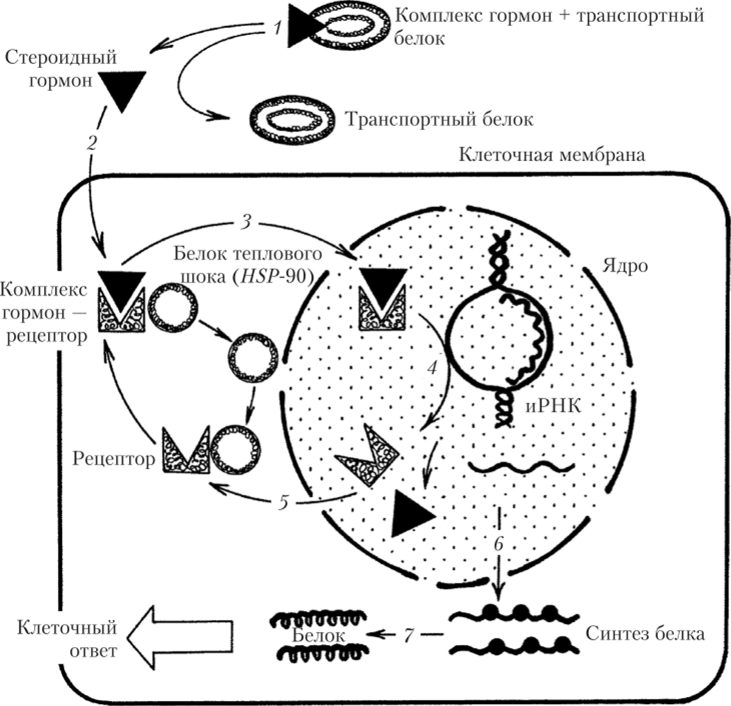
Рис. 1.10. Схема механизма реализации эффектов стероидных гормонов:
- 1 — гормон доставляется к клетке с помощью транспортного белка; 2 — гормон проникает в клетку, диффундируя через мембрану, и образует комплекс с рецептором;
- 3 — от рецептора отделяется белок теплового шока (HSP-90), после чего комплекс гормон — рецептор проходит в ядро; 4 — комплекс гормон — рецептор запускает синтез новой мРНК и распадается; 5 — рецептор выходит из ядра, связывается с белком теплового шока (HSP-90) и многократно используется клеткой;
- 6 — иРНК транспортируется из ядра, и начинается синтез белка рибосомами;
- 7 — синтезированные белки изменяют характер работы клетки
Гормоны, не проникающие через клеточную мембрану, взаимодействуют с рецепторами, расположенными внутри этой мембраны или на ее поверхности.
Некоторые гормоны так же, как и многие медиаторы, способны активировать ионные мембранные каналы, сопряженные с рецепторами или.
являющимися их частью (ионотропные рецепторы). Взаимодействие молекулы такого гормона с рецептором приводит к открытию или, реже, закрытию ионных каналов. Через открытые каналы в клетку проникают ионы, чаще всего Са2+ и (или) Na+, деполяризуя клетку. Ионы кальция являются самыми распространенными вторичными посредниками и мощными регуляторами внутриклеточных процессов. Все это приводит к изменению состояния клетки. Однако значительное число гормонов, для которых характерна мембранная рецепция, не воздействуют на состояние ионных каналов напрямую, а взаимодействуют с рецепторами, сопряженными с G-белками или, реже, с фермент-сопряженными рецепторами (метаботропные рецепторы). Схема сигнализации с помощью ионотропных и метаботропных рецепторов приведена на рис. 1.11.
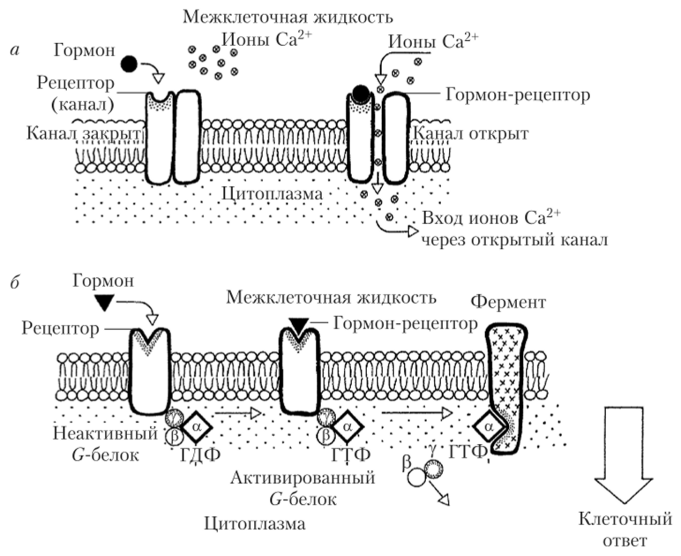
Рис. 1.11. Схема работы гормональных рецепторов:
а — ионотропного (канатыют); б — метаботропного.
Рецепторы, связанные с G-белком, представляют собой аминокислотную последовательность, которая семь раз «прошивает» клеточную мембрану (образуя семь трансмембранных спирализованных участков — доменов). Внеклеточная часть этих белков гликозилирована (рис. 1.12).
Семь трансмембранных доменов расположены в мембране по кругу, образуя в центре углубление со специальными участками связывания лиганда. Присоединение лиганда (в нашем случае — гормона) или агониста приводит к изменению конформации рецепторных белков, благодаря чему активируется G-белок (гуанозинтрифосфат-связывающий белок). G-белок расположен на внутренней стороне мембраны и состоит из трех субъединиц: а, Р и у. Разные G-белки различаются строением а-субъединиц (см. рис. 1.12).
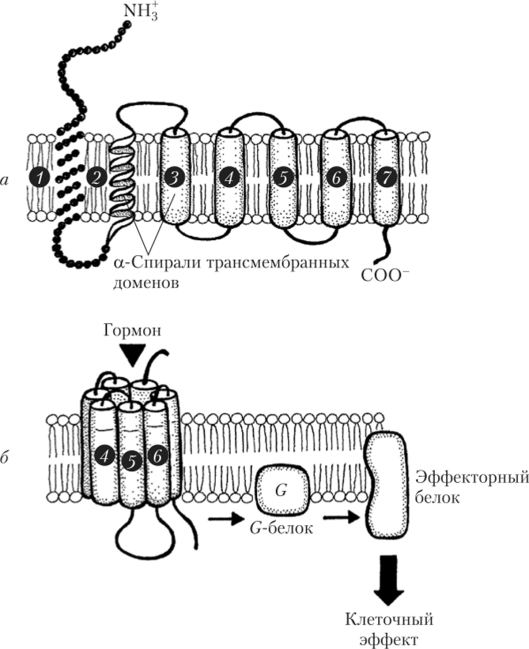
Рис. 1.12. Строение трансмембранных доменов рецепторов (1—7), сопряженных с G-белками (а), и последовательность событий при его гормональной активации (б).
После присоединения гормона к рецептору конформация рецепторного белка изменяется таким образом, что а-субъединица G-белка освобождает ГДФ (гуанозиндифосфат), присоединяет ГТФ (гуанозинтрифосфат) и отделяется от Ру-субъединиц, вступая в контакт с эффекторным белком, изменяя его активность. Ру-Субъединицы также могут оказывать собственные эффекты. а-Субъединица обеспечивает медленный гидролиз связанного ГТФ до ГДФ. Комплекс СЦ-ГДФ не имеет сродства к эффекторным белкам и вновь воссоединяется с Ру-субъединицами. а-Субъединицы G-белков различаются по сродству и типу воздействия на эффектор-
ные белки. Например, Са-ГТФ Gj-белка стимулирует АЦ, в то время как 6'Ц-ГТФ С;-белка ее ингибирует. Са-ГТФ G^-белка активирует фермент фосфолипазу С (рис. 1.13).
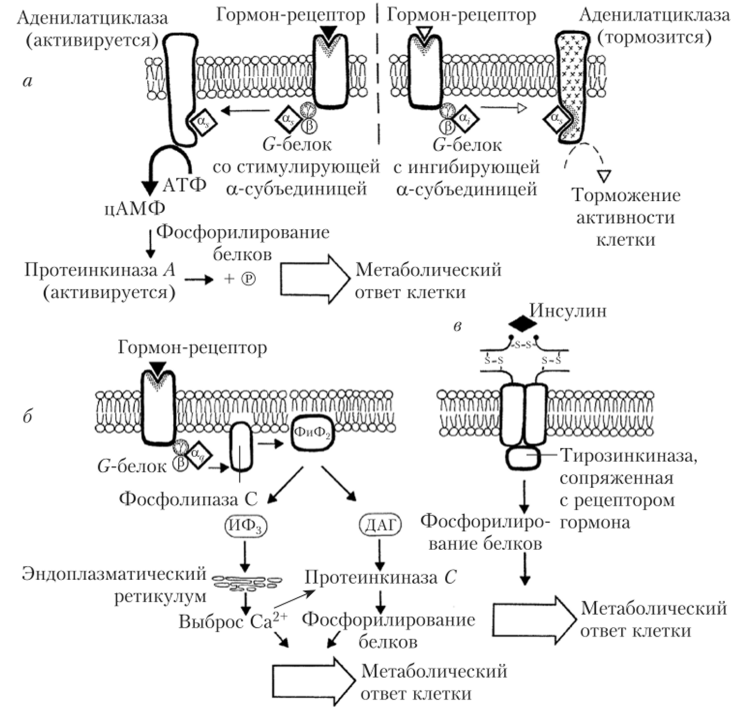
Рис. 1.13. Различные типы метаботропных рецепторов:
а — эффект опосредован через изменение активности АЦ; б — эффект опосредуется через активацию фосфоинозитидной системы; в — эффект опосредуется через активацию тирозинкиназы.
G-белок-связанные рецепторы очень широко представлены в организме. К ним относятся рецепторы к белково-пептидным гормонам, простагландинам, лейкотриенам, НА, дофамину, опиоидам, гистамину и многим другим медиаторам и гормонам. Следует еще раз подчеркнуть, что эти белки отличаются друг от друга только строением а-субъединицы, а в конечном счете — воздействием на разные внутриклеточные регуляторные системы.
В тех случаях, когда G-белок содержит а^-субъединицу, эта субъединица активирует АЦ (рис. 1.13, а). Повышение уровня цАМФ резко повышает уровень жизнедеятельности клетки, так как цАМФ активирует протеинкиназу Ay фосфорилирующую многие регуляторные белки. В частности, фосфорилирование белков Са2+-каналов способствует увеличению поступления Са2+ в клетку. В качестве вторичного посредника систему АЦ — цАМФ используют следующие гормоны:
- — АКТГ;
- — ангиотензин II;
- — кальцитонин;
- — катехоламины (р-рецепторы);
- — кортикотропин-рилизинг-гормон;
- — ФСГ;
- — глюкагон;
- — хорионический гонадотропин человека;
- — ЛГ;
- — паратгормон;
- — секретин;
- — соматостатин;
- — тиреотропный гормон;
- — вазопрессин.
В случае некоторых других гормонов комплекс гормон — рецептор — Gq-белок активирует мембранный фермент фосфолипазу С, которая катализирует гидролиз фосфатидиликозитолдифосфата (ФИФ2) на два важных метаболита, обладающих биологической активностью: инозитолтрифосфат (ИФ3) и диацилглицерол (ДАГ) (рис. 1.13, б). ИФ3 стимулирует выход ионов кальция из внутриклеточных депо через ИФ3-рецепторы, а ДАГ активирует протеинкиназу С, которая фосфорилирует целый ряд белков, изменяя их активность. В целом, наблюдаются высвобождение гормонов, рост клеток, экспрессия генов и т. п. Регуляторный эффект дополняется еще и тем, что липидная часть ДАГ — арахидоновая кислота — является предшественником простагландинов, т. е. гистогормонов, обладающих многими регуляторными эффектами, как на местном тканевом, так и на системном уровнях.
В некоторых клетках G-белки могут прямо или опосредованно воздействовать на молекулы ионных каналов, регулируя ток Са2+ через мембрану. Попав в клетку, ионы кальция связываются с белком кальмодулином. Когда кальмодулин взаимодействует с четырьмя ионами кальция, его конформация меняется, что приводит к активации или ингибированию протеинкиназ, запуская мощный клеточный ответ на воздействие гормона. Например, миозинкиназа действует непосредственно на миозин гладких мышц, вызывая их сокращение.
Целое семейство рецепторов к гормонам содержит встроенные протеинкииазы, которые специфически стимулируют фосфорилирование остатков тирозина в белках-мишенях (рис. 1.13, в). К таким гормонам относятся инсулин, инсулиноподобный фактор роста (ИФР-I) и некоторые другие гормоны.
Еще раз необходимо отметить, что характер воздействия гормона на клетку определяется рецепторами этой клетки. Например, при сахарном диабете только около 30% больных испытывают нехватку инсулина, а у остальных развивается резистентность рецепторов клеток-мишеней к этому гормону. При синдроме тестикулярной феминизации генотип у человека соответствует мужскому (XY), однако фенотипически развивается женщина. У больных происходит мутация двух генов, в которых заложена структура рецепторов к мужскому половому гормону — дигидротестостерону (ДГТ). Этим определяется невозможность для гормона, уровень которого нормален, оказать программирующий эффект, и пол эмбриона остается базовым, т. е. женским. В табл. 1.4 приведены основные типы рецепторов к гормонам и примеры гормонов, взаимодействующих с этими рецепторами.
Таблица 1.4
Основные суперсемейства гормональных рецепторов и взаимодействующие с ними гормоны.
Тип рецептора | Связываемый гормон |
Ядерные | Стероидные гормоны:
|
Тиреоидные гормоны:
| |
Мембранные, сопряженные с G-белками |
|
Мембранные, сопряженные с тирозинкиназами |
|
Мембранные, рецепторные тирозинкипазы и рецепторные гуанилатциклазы |
|