Индивидуальное развитие и эволюция

Недавно (80−90-е гг. XX в.) было показано, что важную роль в контроле процессов онтогенетического формирования пространственной организации тела зародыша играет особая категория генов-регуляторов, именуемых гомеобокссодержащими, или гомеобоксными. Гомеобокс представляет собой эволюционно консервативный генетический блок — последовательность из 180 нуклеотидов, обнаруженный в геноме… Читать ещё >
Индивидуальное развитие и эволюция (реферат, курсовая, диплом, контрольная)
Между мутациями и их проявлением в фенотипе лежат сложные процессы морфогенеза, в ходе которых наследственная информация, преобразуясь, переходйт на все более высокие уровни (молекулы — клетки — ткани — органы). В то же время развивающийся организм подвергается воздействию различных внешних факторов, направляющих процессы морфогенеза в те или другие русла, приводя к формированию конкретных фенотипических особенностей. Сущность этих процессов и механизмы преобразования информации известны еще сравнительно слабо; обобщающая теория индивидуального развития еще не создана. Это очень затрудняет использование эмбриологических концепций в эволюционистике. Данные и обобщения онтогенетики еще не заняли подобающего места в синтетической теории эволюции.
Дифференцировка организма в онтогенезе
Одной из основных проблем онтогенетики, решение которой должно сыграть важную роль и для эволюционной теории, остается сущность явлений онтогенетической дифференцировки, т. е. тех процессов, которые ведут к развитию разнокачественности различных фенотипических структур, возникающих в онтогенезе, в конечном итоге, из единственной клетки — зиготы. Даже в организации столь просто устроенных животных, как кишечнополостные, насчитывается около 10 разных типов клеток, а в организме млекопитающих — порядка 1000 типов клеток. Клетки разных типов группируются в ткани и формируют многотканевые комплексы со сложной пространственной структурой — органы.
По современным представлениям, в процессе дробления зиготы на бластомеры и при последующих делениях соматических клеток каждая дочерняя клетка в норме получает 100% наследственной информации, т. е. полный набор генов, точно соответствующий исходному генотипу зиготы. Однако соматические клетки резко различаются по строению и функциям (достаточно сравнить, например, нейрон, мышечную клетку и эритроцит в одном организме).
Можно считать установленным, что различия соматических клеток определяются дифференциальной активностью генов (другими словами, в одних типах клеток активны одни, а в других — другие участки хромосом). Неактивные в данной клетке гены блокируются какими-то входящими в состав хромосом веществами, какими именно — еще точно не выяснено. Предполагают, что этими веществами (репрессорами) являются те или иные хромосомные белки (многие ученые считают репрессорами белки-гистоны).
Очевидно, при эквивалентности генома всех зародышевых клеток их первоначальная дифференцировка должна определяться разнокачественностью их цитоплазмы. В период дробления практически не происходит роста клеток между их последовательными делениями. Поэтому в разные бластомеры попадают различные и, вероятно, разнокачественные участки цитоплазмы зиготы. Возможно, разнокачественность цитоплазмы и определяет активацию различных генов в разных бластомерах. Деятельность активированных генов приводит к изменениям цитоплазмы бластомеров — специфическим в разных клетках. Измененное цитоплазматическое окружение взаимодействует с геномом клетки, вызывая избирательную активацию новых групп генов и репрессируя другие группы. Так запускается цепь последовательных взаимодействий между геномом и цитоплазмой, протекающая по принципу обратной связи (положительной для одних и отрицательной для других генных комплексов), в результате которых клетка дифференцируется в определенном направлении (рис. 99). Однако пока еше мало известно о конкретных взаимодействиях между ядром и цитоплазмой на разных этапах клеточной дифференцировки.
Онтогенез отнюдь не сводится.
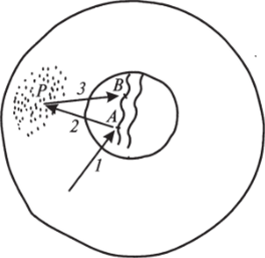
Рис. 99. Схема клеточной дифференцировки:
- 1 — активация цитоплазматическим белком локуса, А в хромосоме ядра;
- 2 — трансляция иРНК, транскрибированной на активированном локусе, на рибосомы (Р); 3 — белок, синтезированный на рибосомах, поступает в цитоплазму и в ядро, где
активирует новый локус (В) к дифференциации разных типов клеток. Важнейшим аспектом индивидуального развития многоклеточных организмов является установление общего плана их строения, т. е. оформление пространственной структуры организма и определенного расположения его частей. У всех более сложных многоклеточных организмов к этому добавляются процессы органогенезов — формирование дифференцированных органов. Все эти процессы осуществляются посредством взаимодействия друг с другом отдельных клеток и клеточных комплексов.
О сущности этих взаимодействий известно, пожалуй, еще меньше, чем о молекулярных механизмах клеточной дифференцировки, хотя исследования в этой области ведутся уже давно. Одной из наиболее разработанных концепций является теория эмбриональной индукции, созданная трудами Г. Шпемана и его учеников.
Под эмбриональной индукцией понимается такое взаимодействие между двумя клеточными комплексами зародыша, при котором один из них — организатор или организационный центр — направляет (индуцирует) дифференциацию другого — реагирующей ткани — в определенном направлении. В нормальных условиях, т. е. в живом зародыше, присутствие организатора необходимо для формирования из реагирующих тканей специфических структур взрослого организма. Так, у зародышей позвоночных крыша первичной кишки, образованная зачатком хорды, индуцирует в расположенной над ней эктодерме развитие нервной системы; глазной бокал, растущий по направлению к покровам от зародышевого головного мозга, индуцирует развитие хрусталика глаза; зачаток внутреннего уха (слуховой пузырек) индуцирует развитие вокруг себя слуховой капсулы и т. п. Согласно теории эмбриональной индукции, дифференциация органов в индивидуальном развитии представляет собой цепь индукционных взаимодействий, в которую последовательно включаются новые и новые организаторы, и каждый предшествующий этап «запускает в действие» следующий.
Характер взаимодействий между морфогенетическими системами в процессах эмбриональной индукции может изменяться в филогенезе. Например, у эмбрионов ряда костных рыб (Amia, Salmo) невромасты сейсмосенсорных рецепторов (или органов боковой линии) в области головы играют роль индукторов для формирования накладных костей черепа. Однако у хрящевых рыб невромасты сейсмосенсорных рецепторов в области головы имеются, но накладные кости черепа не развиваются. Следовательно, индукционные взаимодействия здесь разорваны: либо невромасты потеряли свойства индукторов, либо реагирующие ткани утратили свою реактивность. В то же время у наземных позвоночных накладные кости черепа имеются, а органов боковой линии нет. Очевидно, в этом случае вместо старого механизма индукции возник новый. С другой стороны, некоторые индукционные системы поразительно консервативны, сохраняясь у очень далеких друг от друга групп — таков первичный хордомезодермальный организатор у всех хордовых.
Природа эмбриональной индукции еще не вполне выяснена. Индукция может основываться на непосредственном взаимодействии поверхностных мембран клеток организатора и реагирующей ткани, или на химической передаче индуцирующего действия, или на совокупности пространственных и химических взаимодействий. По некоторым данным, в химической индукции существенную роль может играть белковый компонент рибонуклеопротеидов индуктора.
Явление эмбриональной индукции, несомненно, играет важную роль в онтогенезе, но отнюдь не исчерпывает всех форм взаимодействий между клеточными комплексами зародыша, обеспечивающих развитие структурной упорядоченности организма[1].
Недавно (80−90-е гг. XX в.) было показано, что важную роль в контроле процессов онтогенетического формирования пространственной организации тела зародыша играет особая категория генов-регуляторов, именуемых гомеобокссодержащими, или гомеобоксными. Гомеобокс представляет собой эволюционно консервативный генетический блок — последовательность из 180 нуклеотидов, обнаруженный в геноме филогенетически столь далеких друг от друга организмов, как позвоночные, иглокожие, насекомые и нематоды. Гомеобокс кодирует «гомеодомен» из 60 аминокислот. Протеины, содержащие этот гомеодомен, регулируют транскрипцию других генов. Гомеобоксные гены являются важными регуляторами, контролирующими онтогенетические программы, которые определяют формирование специфики разных сегментов вдоль передне-задней оси тела. Очевидно, мутации гомеобоксных генов могут иметь очень глубокие последствия для организма, приводя к изменениям соответствующих программ развития. Значительное большинство исследованных мутаций гомеобоксных генов (например, у мух-дрозофил) летальны, что вполне естественно для крупных мутаций. Эти обстоятельства определяют высокий эволюционный консерватизм гомеобоксных генов, сохраняющих значительную степень общности у филогенетически далеко разошедшихся групп организмов.
- [1] См.: Т о к и н Б. П. Общая эмбриология. — М., 1970; Белоусов Л. В.
Введение
в общую эмбриологию. — М., 1980.