Пищеварение в желудке

Кишечная фаза желудочной секреции начинается после запуска эвакуации пищевых масс из желудка. Продукты гидролиза белков, поступившие в двенадцатиперстную кишку, активируют ее хемои механорецепторы, включая местные (метасимпатические) тормозные рефлексы. Их дуги замыкаются с участием нейронов межмышечного нервного сплетения стенки пищеварительного тракта, результатом чего становятся ослабление… Читать ещё >
Пищеварение в желудке (реферат, курсовая, диплом, контрольная)
Пищеварение в желудке представляет собой ряд взаимосвязанных процессов:
- — накопление и хранение пищи — за счет растяжимости стенки;
- — механическая переработка нищи — благодаря развитой моторике;
- — стерилизация пищевой массы под действием НС1;
- — химическая (НС1) и ферментативная (пепсины, липаза) переработка пищи;
- — всасывание — в желудке происходит всасывание воды, солей, глюкозы и ряда других, в том числе лекарственных, веществ;
- — выработка гастроинтестинальных гормонов, в том числе гастрина, серотонина, мотилина и ряда других;
выработка регуляторных факторов, таких как внутренний фактор Кастла;
— выделение балластных продуктов обмена — аммиака, мочевой кислоты, мочевины, креатина.
В зависимости от количества и химического состава пища находится в желудке от 2 до 10 ч. Все это время она измельчается, разжижается, перемешивается и пропитывается желудочным соком, с помощью которого осуществляется переваривание.
Желудок представляет собой толстостенное расширение пищеварительного тракта мешкообразной формы, расположенное в брюшной полости (рис. 5.14).
Место сообщения желудка с пищеводом называется кардиальным отделом. Центральная часть желудка называется телом или фундальным отделом. Место сообщения желудка с двенадцатиперстной кишкой называется пилорическим отделом. В этом отделе располагается пилорический сфинктер (привратник). Изнутри стенка желудка имеет многочисленные складки — кристы, которые значительно увеличивают площадь ее соприкосновения с пищей и способствуют механической обработке пищи.
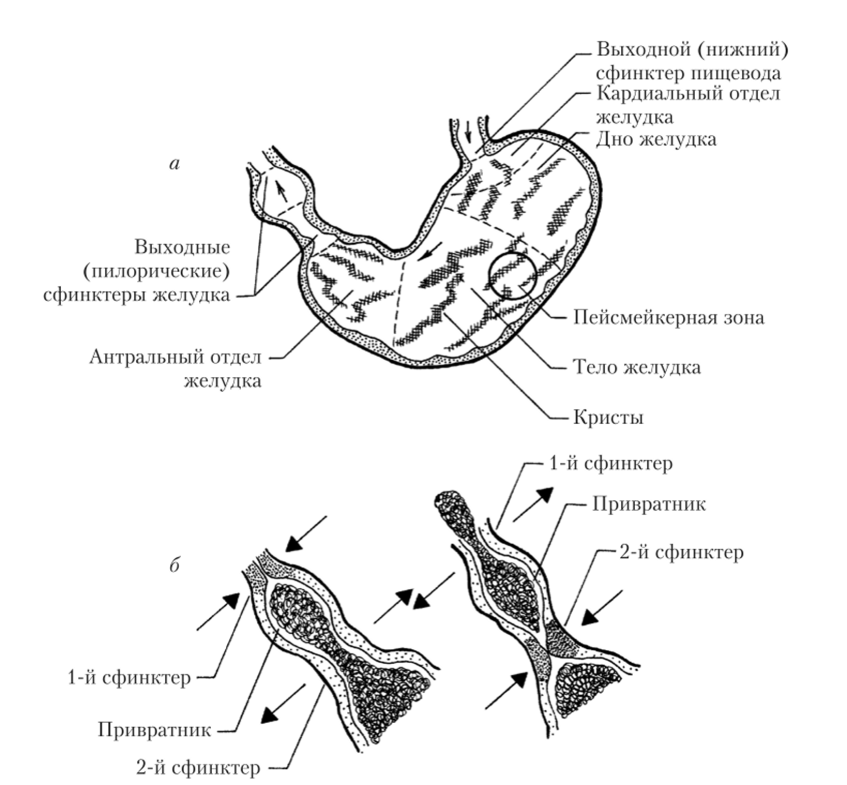
Рис. 5.14. Основные структурные и функциональные области желудка (а) и схема работы его выходных сфинктеров (б)
В разных отделах желудка располагаются многочисленные простые железы, различающиеся как по строению, так и по составу продуцируемого секрета. В кардиальном отделе лежат простые трубчатые сильно разветвленные железы, вырабатывающие, главным образом, слизь и бикарбонаты (НС03), и совсем мало ферментов. В пилорическом отделе желудка лежат простые трубчатые слаборазвитые железы, также секретирующие слизь, бикарбонаты и небольшое количество ферментов, но отличающиеся большой эндокринной активностью. Простые трубчатые неразветвленные железы лежат в фундальном отделе. Именно они секретируют основное количество ферментов желудочного сока и соляную кислоту (НС1), а также слизь, бикарбонаты и гормоны, но в значительно меньшем количестве (рис. 5.15).
Протоки желез выводят секрет в небольшие углубления слизистой оболочки желудка, которые называются желудочными ямками. Совокупность секретов пищеварительных желез желудка называется желудочным соком.
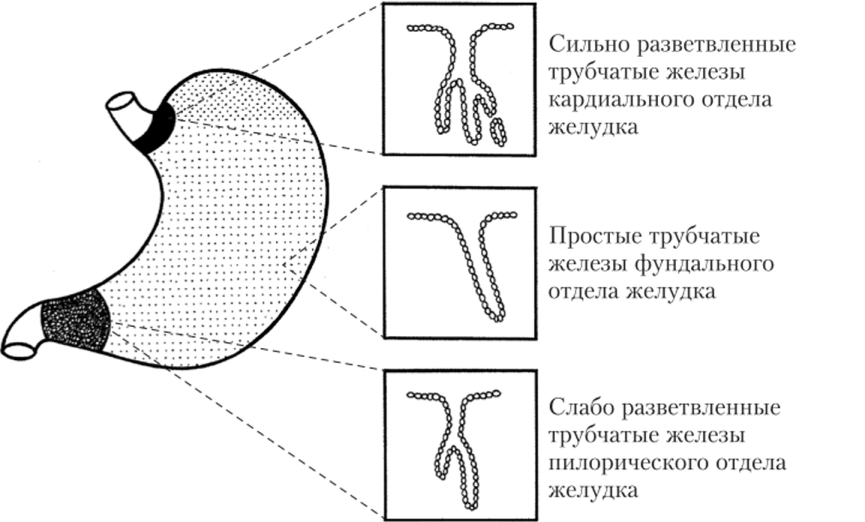
Рис. 5.15. Расположение разных типов трубчатых желез в отделах желудка.
Желудочный сок вырабатывается железами слизистой оболочки желудка с большой интенсивностью: за сутки образуется от 2 до 2,5 л.
Неферментативная часть желудочного сока включает НС1 (0,3—0,5%), неорганические ионы (Na+, К+, Са2+, СП, НСО3), а также слизь и конечные продукты метаболизма. Уровень pH чистого желудочного сока составляет 1,5−1,8.
Ферментативная часть желудочного сока представлена в основном протеолитическими ферментами — пенсинами: пепсином А, гасгриксином и пепсином В (желатиназой). Эти ферменты вырабатываются и сскрстируются в неактивной форме — в виде пепсиногенов. В дальнейшем под влиянием соляной кислоты от пепсиногенов отщепляются небольшие фрагменты, что превращает их в активные пепсины. Такой механизм позволяет защитить секреторные клетки от повреждения собственными ферментами. Пепсины расщепляют белки пищи на пептидные фрагменты и отдельные аминокислоты. Разные пепсины действуют на разные белки и при различных значениях pH, обеспечивая, в числе прочего, всеядность человека.
Помимо пепсинов, в состав желудочного сока входит небольшое количество липазы. Она способна расщеплять эмульгированные жиры, например жир в составе молока. Активность желудочной липазы особенно велика в случае грудных детей, у которых она расщепляет более половины жиров, входящих в состав материнского молока.
Функции соляной кислоты в желудке важны и многообразны. Она активирует пепсиногены, превращая их в пепсины и одновременно создавая кислую среду, оптимальную для протеолиза. Соляная кислота вызывает денатурацию белков пищи, что делает их более доступными для расщепления ферментами. Кроме того, создаваемый НС1 уровень кислотности влияет на моторику стенок желудка и выработку желудочного сока. Так, преждевременное поступление в двенадцатиперстную кишку слишком кислого химуса тормозит сокращения желудка, а снижение pH ниже 3,0 ослабляет секрецию желудочного сока. Важно и то, что соляная кислота обладает бактерицидным действием.
Функции слизи в желудке обусловлены тем, что она вместе с ионами НСО3 образует вязкую гидрофобную массу (гель), которая защищает стенки желудка от повреждающего действия НС1 и пепсинов. Слизь, которую образуют железы нижней части желудка, содержит, помимо гликопротеина муцина, особый белок — внутренний фактор Кастла. Этот белок специфически связывает витамин В12, поступающий с пищей, защищает его от разрушения и способствует всасыванию в тонком кишечнике. Витамин В12 играет важную роль в кроветворении, регулируя созревание клеток — предшественников эритроцитов (эритропоэз). Дефицит этого витамина вызывает тяжелую форму анемии.
Расположение секреторных клеток в трубчатой железе всегда соответствует определенным правилам:
- — добавочные клетки, вырабатывающие слизь и бикарбонаты, всегда располагаются около выхода из железы;
- — клетки, вырабатывающие соляную кислоту (обкладочные, или париетальные), расположены несколько глубже добавочных;
- — главные клетки, секретирующие пепсиноген и липазу, располагаются глубже добавочных и обкладочных;
- — на дне железы располагаются клетки, секретирующие гастроинтестинальный гормон гастрин (G-клетки);
- — клетки, секретирующие гистамин и соматостатин, лежат в глубине слизистой оболочки, примыкая к эпителиальным секреторным клеткам (рис. 5.16).
Секреция соляной кислоты осуществляется обкладочными (париетальными) клетками. Особенностями их строения является наличие системы особых внутриклеточных канальцев. Увеличение активности обкладочных клеток приводит к открытию этих канальцев, тянущихся в глубину клетки из просвета железы. Многочисленные тубуловезикулы, примыкающие изнутри к мембранам канальцев, сливаются с ними, образуя складки (микроворсинки). Эти складки сильно увеличивают поверхность мембраны, в которую встроено большое количество К+— и С1″ -каналов, а также молекул К+/Н+-АТФ-азы (рис. 5.17).
К+/н+_АТФ-аза в мембране каналов обкладочных клеток переносит протоны Н+ в обмен на катионы К+. Ионы К+ возвращаются назад через К+-каналы. На каждый протон, вышедший в каналец, один бикарбонатанион (НСО3) проходит через мембрану базальной части клетки в тканевую жидкость, обмениваясь с помощью анионного транспортера на анион С1~. Бикарбонат-анионы НСО3 образуются в ходе катализируемой карбоангидразой реакции:
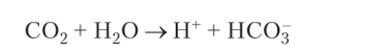
В результате этих процессов в клетке накапливается избыток анионов С1, которые по хлорным каналам выходят в просвет внутриклеточных канальцев (рис. 5.18).
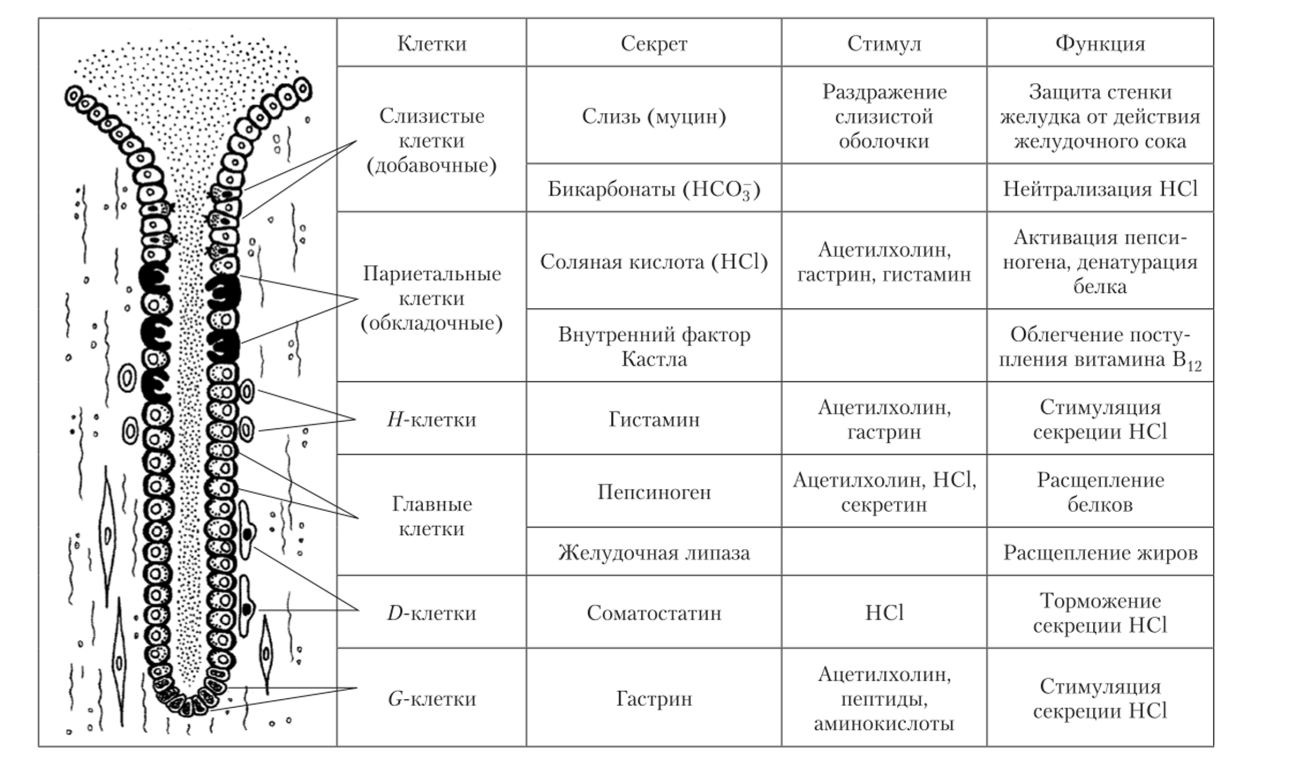
Рис. 5.16. Основные секреторные клетки трубчатых желез желудка, их секрет, стимулы к активации и функции.
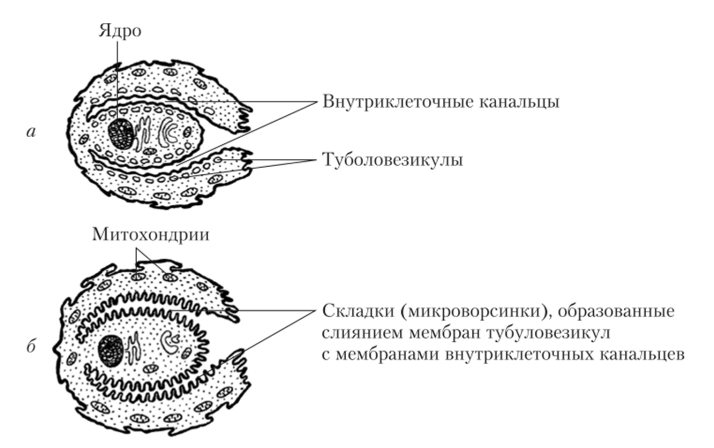
Рис. 5.17. Морфологические изменения в париетальных клетках при усилении их секреторной активности:
а — невысокая секреторная активность; б — высокая секреторная активность.
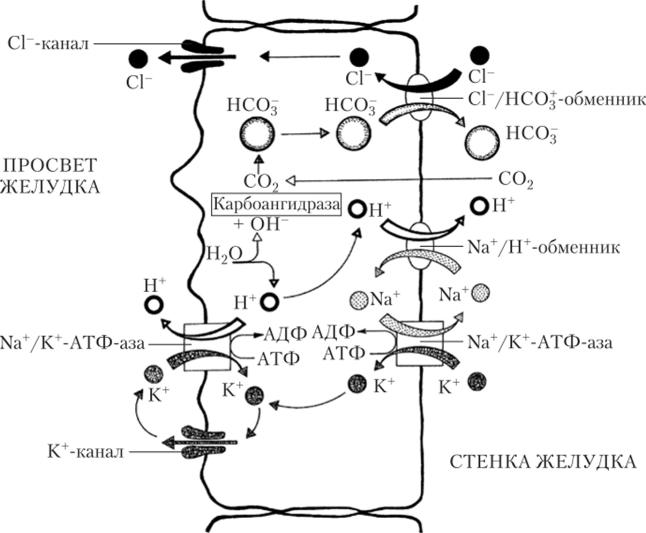
Рис. 5.18. Механизм секреции компонентов соляной кислоты (HCl) париетальными клетками желез желудка.
Секреция прогонов Н+, в покое составляющая около 2 ммоль/ч, в процессе пищеварения может возрастать до 20 ммоль/ч.
Двигательная активность желудка и механизмы ее регуляции
сильно различаются в двух его отделах: верхнем и нижнем.
Мышцы стенки верхнего отдела постоянно находятся в равномерном тоническом напряжении. Тонус зависит от эфферентных преганглионарных волокон, которые приходят в желудок в составе блуждающего нерва и образуют синапсы на постганглионарных холинэргических нейронах межмышечного сплетения. При приеме пищи тонус стенок верхнего отдела снижается, желудок расширяется и наполняется, вмещая (депонируя) до 3 л пищевой массы. Рефлекс релаксации запускается механорецепторами глотки и пищевода, а рефлекс расширения — механорецепторами стенки самого желудка, сигналы от которых направляются в продолговатый мозг. Эфферентные преганглионарные волокна этих двух рефлексов приходят в желудок в составе блуждающего нерва и образуют синапсы на тормозных метасимпатических нейронах межмышечного сплетения.
Нижний отдел желудка участвует в перемешивании и гомогенизации пищи, а также ее перемещении в двенадцатиперстную кишку. Мембранный потенциал гладкомышечных клеток этого отдела ритмически изменяется (медленные волны). Источником таких спонтанных колебаний является пейсмейкерная зона, располагающаяся в межмышечном сплетении средней части желудка (рис. 5.14, а). Некоторые из колебаний переходят в ПД, обусловленные входом в клетки, прежде всего, ионов кальция (см. рис. 5.6). В результате возникают циклы перистальтических сокращений, которые распространяются по желудку в направлении пилорического сфинктера.
Интенсивность перистальтики желудка находится под нервным и гуморальным контролем. Расширение верхнего отдела желудка стимулирует перистальтику нижнего отдела. Этот рефлекс запускается механорецепторами стенки верхнего отдела, сигналы от которых активируют в продолговатом мозге парасимпатические преганглионарные нейроны. Их аксоны в составе блуждающего нерва идут к возбуждающим нейронам межмышечного сплетения.
Растяжение тонкого кишечника тормозит перистальтику нижнего отдела желудка. Соответствующий рефлекс запускается механорецепторами тонкого кишечника, замыкается в продолговатом мозге, а реализуется симпатическими волокнами, которые тормозят возбуждающие нейроны межмышечного сплетения.
Регуляция желудочной секреции включает три фазы: цефалическую, желудочную и кишечную.
Цефалическая фаза начинается еще до попадания пищи в ротовую полость. Обстановка, вид и запах пищи действуют как условные раздражители на рецепторы органов чувств. Попадание пищи в ротовую полость активирует тактильные, температурные и вкусовые рецепторы слизистой оболочки полости рта, глотки и пищевода. В результате активируется комплекс условных и безусловных рефлексов, дуги которых замыкаются на ядрах блуждающих нервов в продолговатом мозге. Эфферентные волокна блуждающего нерва достигают стенки желудка, стимулируя секрецию желудочного сока (рис. 5.19).
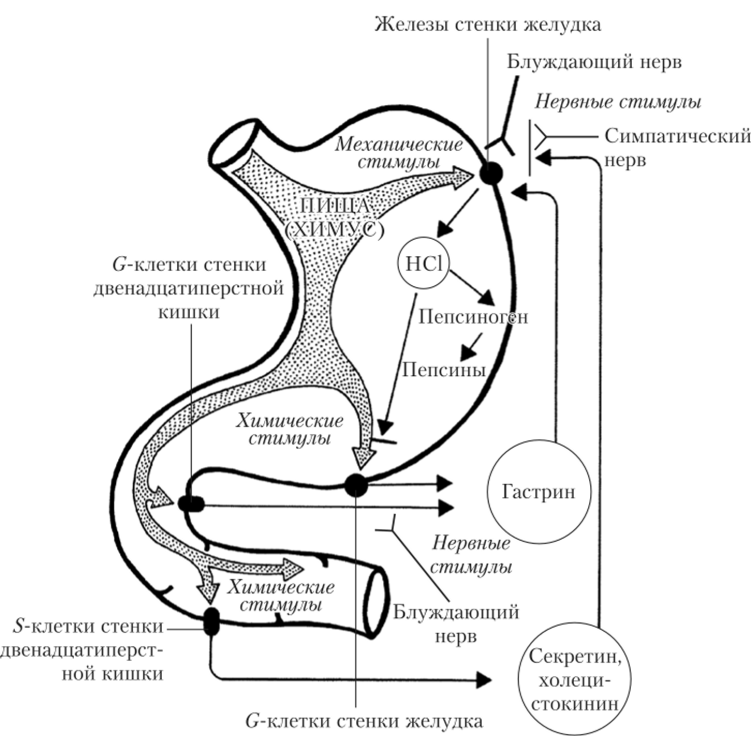
Рис. 5.19. Пути влияния на секрецию компонентов желудочного сока механических и химических стимулов (со стороны пищи) и нервных стимулов (со стороны парасимпатического отдела НС):
|$ZZ — тормозные нервные и химические стимулы;
$ZZ — возбуждающие нервные и химические стимулы Желудочная секреция в эту фазу легко тормозится посторонними раздражителями (эмоциональными, болевыми и др.), что негативно влияет на пищеварение. Тормозные влияния осуществляются с помощью симпатических нервов.
Желудочная, или нейрогуморальная, фаза секреции начинается с попадания пищи в желудок. Возникающая при этом активация механо-, хемои терморецепторов слизистой оболочки рефлекторно активирует главные и обкладочные клетки желудочных желез. Нервная регуляция секреции при этом дополняется гуморальной. В слизистой оболочке нижнего отдела желудка располагаются G-клетки, вырабатывающие гормон гастрин. Гормон выбрасывается в кровь в ответ на механическое растяжение желудка пищей, на воздействие продуктов переваривания белков, а также возбуждение блуждающих нервов (см. рис. 5.19).
Гастрин действует эндокринным путем на обкладочные клетки желудочных желез, усиливая секрецию компонентов соляной кислоты. Помимо обкладочных клеток, гастрин активирует особые клетки желудка, производящие гистамин (//-клетки). Гистамин не поступает в кровь, а дополнительно стимулирует продукцию соляной кислоты лежащими рядом обкладочными клетками (паракринное действие). Выброс гистамина усиливается также при парасимпатических нервных воздействиях (рис. 5.20).
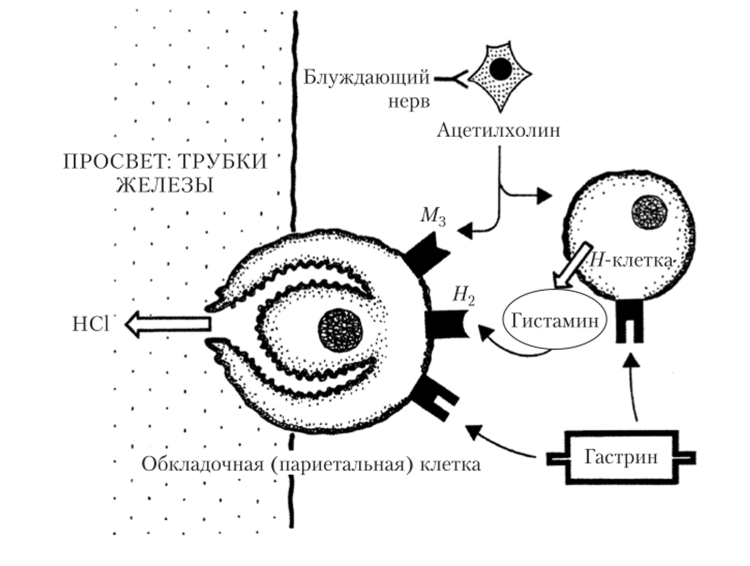
Рис. 5.20. Механизм влияния гастрина и активности парасимпатического отдела ВНС на секрецию соляной кислоты (НС1) париетальными клетками трубчатых желез стенки желудка Главные клетки желудочных желез также являются мишенью гуморальных регуляторных влияний, ведущая роль в которых принадлежит соляной кислоте, оказывающей стимулирующее действие.
Секреция желудка в нейрогуморальную фазу зависит от состава пищи. При длительном употреблении преимущественно углеводной пищи выделение желудочного сока снижается, а при длительном употреблении белковой пищи — возрастает. Наличие в пище острых и экстрактивных веществ также стимулирует работу желудочных желез.
Кишечная фаза желудочной секреции начинается после запуска эвакуации пищевых масс из желудка. Продукты гидролиза белков, поступившие в двенадцатиперстную кишку, активируют ее хемои механорецепторы, включая местные (метасимпатические) тормозные рефлексы. Их дуги замыкаются с участием нейронов межмышечного нервного сплетения стенки пищеварительного тракта, результатом чего становятся ослабление и прекращение желудочной секреции. Одновременно с развитием этих рефлексов идут гуморальные взаимодействия желудка и кишечника. Кислое содержимое желудка (pH менее 3,0) стимулирует выработку стенкой двенадцатиперстной кишки гормона секретина, а продукты переваривания белков и жиров — гормона холецистокинина. Секретин и холецистокинин тормозят работу обкладочных клеток желудка (см. рис. 5.19). Объектом нервной и гуморальной регуляции в стенке двенадцатиперстной кишки, значимой для желудка, являются G-клетки, вырабатывающие гастрип. Аминокислоты химуса стимулируют их работу, а высокая кислотность (pH ниже 3,0) — тормозит. Помимо прямого торможения G-клеток, высокая кислотность стимулирует выработку D-клетками гормона соматостатина. Активность блуждающего нерва влияет на G-клетки через нейроны энтеральной НС. Стимулирующие влияния опосредуются гастрин-высвобождающим пептидом, а тормозные — через активацию D-клеток АХ (рис. 5.21).
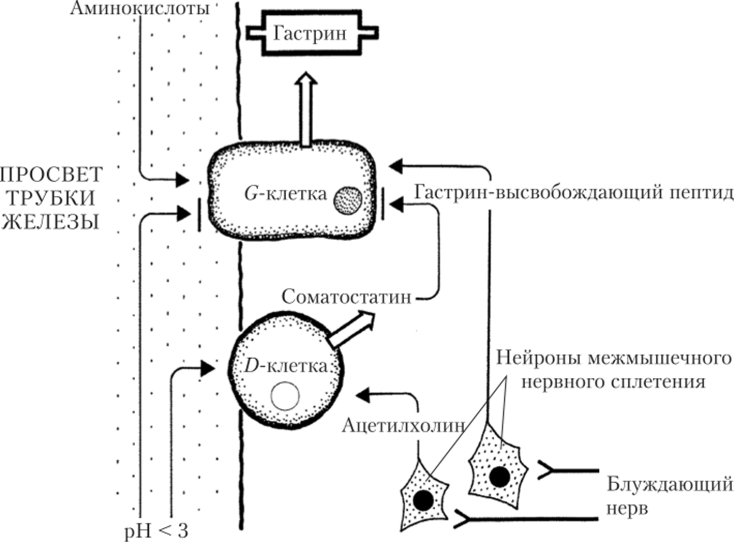
Рис. 5.21. Механизм регуляции секреции гастрина G-клетками в стенке двенадцатиперстной кишки.