Особенности использования многоуглеродных субстратов микроорганизмами

У многих аэробных и анаэробных почвенных и фитопатогенных бактерий (цитофаг, миксобактерий, клостридий, эрвиний) и грибов имеются экзоферменты для гидролиза целлюлозы, ксилана древесины и луба, крахмала, пектина, агара, хитина, лигнина. Образовавшиеся при гидролизе дисахариды далее подвергаются либо гидролизу, либо фосфоролизу. Моносахара превращаются в пируват по одному из рассмотренных ранее… Читать ещё >
Особенности использования многоуглеродных субстратов микроорганизмами (реферат, курсовая, диплом, контрольная)
Окисление многоуглеродного субстрата начинается с его преобразования в вещество, которое клетка способна транспортировать внутрь и с которым смогут работать клеточные ферменты. У микроорганизмов подготовительными реакциями использования биополимеров будут реакции гидролиза с помощью выделяемых в среду ферментов. Так, для окисления белков некоторые патогенные, портящие продукты, почвенные бактерии и грибы синтезируют внеклеточные протеазы, которые гидролизуют белки до коротких пептидов и аминокислот. Разложение белка микроорганизмами (аммонификация) всегда сопровождается образованием ряда продуктов: аммиака при дезаминировании аминокислот, сероводорода при расщеплении серосодержащих аминокислот (метионина, цистеина и их производных) и индола при распаде триптофана. При этом происходит подщелачивание среды. Аминокислоты дезаминируются с образованием органических кислот (пирувата, ацетата и интермедиатов ЦТК) и в виде ацетил-КоА входят в цикл Кребса для полного окисления и получения клеткой энергии. Также они могут быть использованы как источник углерода для синтезов клеточных компонентов. Аминогруппы аминокислот идут либо на анаболизм азотсодержащих веществ клетки, либо выделяются в виде иона аммония.
У многих аэробных и анаэробных почвенных и фитопатогенных бактерий (цитофаг, миксобактерий, клостридий, эрвиний) и грибов имеются экзоферменты для гидролиза целлюлозы, ксилана древесины и луба, крахмала, пектина, агара, хитина, лигнина. Образовавшиеся при гидролизе дисахариды далее подвергаются либо гидролизу, либо фосфоролизу. Моносахара превращаются в пируват по одному из рассмотренных ранее путей: гликолизу, пентозофосфатному или КДФГ-пути. В аэробных условиях ацетил-КоА образуется из пирувата с помощью пируватдегидрогеназного комплекса с тиамином (Bt) в качестве кофермента.
Липиды расщепляются микробными липазами до глицерола и жирных кислот. Глицерол затем фосфорилируется, окисляется до ДОАФ и катаболизируется через реакции гликолиза. Жирные кислоты подвергаются р-окислению до ацетил-КоА, метаболизируемого в цикле Кребса.
Ароматические соединения сначала подвергаются атаке моноили диоксигеназ, внедряющих в бензольное кольцо один или два атома кислорода, соответственно. Образовавшаяся р-кетоадипиновая кислота или ее производное затем расщепляется до линейного продукта, который разлагается на ацетил-КоА и сукцинат, окисляющиеся в ЦТК.
Предельные и непредельные углеводороды (кроме одноуглеродных соединений) используются почвенными микроорганизмами разных групп. Сначала они окисляются до соответствующих жирных кислот с помощью оксигеназ, а затем подвергаются Р-окислению.
Производные пуринов и пиримидинов «входят» в ЦТК через фумарат, а пентозы — через а-кетоглутарат.
Многоуглеродные субстраты могут при дыхании подвергаться полному или неполному окислению. При полном окислении единственным окисленным продуктом будет С02, а конечным этапом окисления, как правило, цикл Кребса. В качестве альтернативных путей конечного окисления следует назвать цикл дикарбоновых кислот у Е. coli (рис. 7.25) и окислительный пентозофосфатный путь (Варбурга — Диккенса — Хорекера) у бактерий рода Gluconobacter. В последнем случае, чтобы молекула гексозы окислилась полностью, цикл должен «прокрутиться» шесть раз.
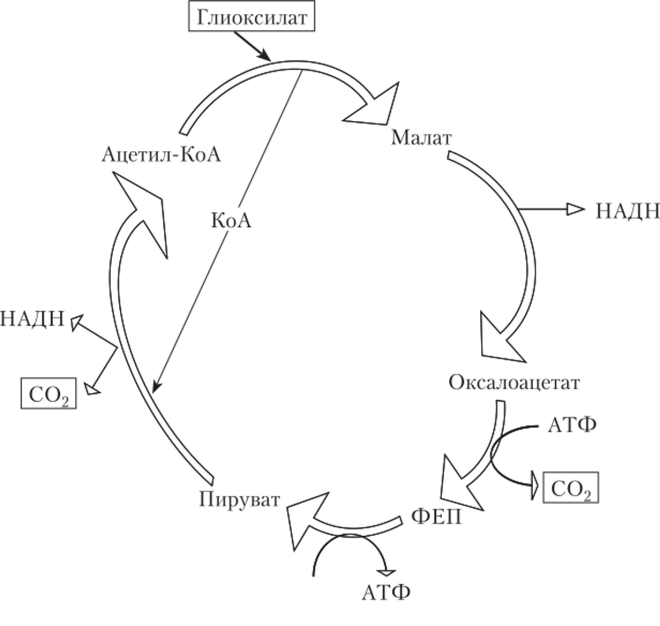
Рис. 7.25. Цикл дикарбоновых кислот у Е. coli.
При неполном окислении питательного субстрата в качестве конечных продуктов в среду выделяются наряду с углекислым газом различные органические соединения, часто схожие с продуктами некоторых брожений. Неполное окисление многоуглеродных органических субстратов происходит либо из-за дефектов ферментных систем микроорганизмов, либо в результате существенного замедления их работы из-за неоптимальности условий культивирования. Типичными представителями микроорганизмов, осуществляющих неполные окисления, являются уксуснокислые бактерии. Эти грамотрицательные мелкие палочки широко распространены в природе и живут на листьях растений. Они устойчивы к кислотам (до pH = 3—5) и к этанолу (до 9—10%). Уксуснокислые бактерии способны образовывать органические кислоты путем неполного окисления сахаров и спиртов. Наиболее известны два рода: Acetobacter с перитрихиальным жгутикованием и Gluconobacter с полярно расположенными жгутиками. У представителей рода Acetobacter высокая активность алкогольдегидрогеназы сочетается с низкой активностью ферментов ЦТК, а у бактерий рода Gluconobacter отсутствует ключевой фермент ЦТК а-кетоглутаратдегидрогеназа. Уксуснокислые бактерии используют в биотехнологии для превращения этанола в ацетат, сорбита в сорбозу, образования глюконата и ацетона.
Многие грибы способны выделять в среду продукты метаболизма (органические кислоты и аминокислоты). Эго связано с «дезорганизацией» их метаболизма путем изменения условий культивирования, затормаживающих деятельность определенных ферментов ЦТК. Причиной накопления промежуточных продуктов в «узких местах» может быть избыток субстрата, изменение pH и концентрации микроэлементов. С помощью грибов можно получать (в том числе и в промышленных масштабах) следующие продукты: молочную (Rhizopus nigricans, Saprolegnia), фумаровую (Mucor, Rhizopus), глюконовую {Aspergillus niger), щавелевую (многие грибы при высоком pH), лимонную (Л. niger) и итаконовую (Л. itaconicus, A. terreus) кислоты. L-глутаминовая кислота образуется промышленным продуцентом Corynebacterium glutamicum путем восстановительного аминирования накапливающегося интермедиата цикла Кребса — а-кетоглутаровой кислоты.