Мембранный потенциал клетки, или потенциал покоя

Бернштейн предложил использовать приведенное выше уравнение Нернста для определения мембранного потенциала, как только узнал, что именно К+ может проходить через мембрану покоящейся возбудимой клетки. Однако соотношение концентраций катионов калия внутри и снаружи клетки оказалось не 1: 10 (как в искусственной экспериментальной системе Нернста), а совсем другим. Так, в мышечных клетках калия в 49… Читать ещё >
Мембранный потенциал клетки, или потенциал покоя (реферат, курсовая, диплом, контрольная)
Возбудимость — способность клеток и тканей переходить в состояние физиологической активности иод влиянием каких-либо внешних воздействий (раздражителей), достигших пороговой величины. Возбудимостью обладают любые живые клетки, в том числе растительные, но в наибольшей степени она выражена у клеток животных — таких, как нервные, мышечные и железистые.
Любому специалисту, связанному с исследованиями организма животного и человека, необходимо понимать, что именно возбудимые клетки составляют основу НС, воспринимают сигналы из внешней и внутренней среды организма, обеспечивают его ответные реакции.
Все возбудимые клетки обладают тремя свойствами:
1) ПН, или мембранным потенциалом, — разностью электрического потенциала между внутриклеточной средой и средой, окружающей клетку;
- 2) IIД — способностью генерировать краткий электрический ответ под действием любого раздражителя, интенсивность которого достигла определенного порога;
- 3) проводимостью — способностью распространять ПД по телу клетки и ее отросткам.
Проведем следующий эксперимент. Если взять нейрон, находящийся в настоящий момент в состоянии покоя, и ввести тонкий стеклянный микроэлектрод с кончиком диаметром не более 1 мкм, то такой кончик не нанесет клетке видимых повреждений. Полость стеклянного электрода должна быть заполнена жидкостью, хорошо проводящей ток (электролитом). Чаще всего для этой цели используют раствор хлористого калия (КС1). Электрод соединяют с вольтметром. Пока кончик электрода находится в межклеточной среде, стрелка микровольтметра стоит на нуле (рис. 8.1).
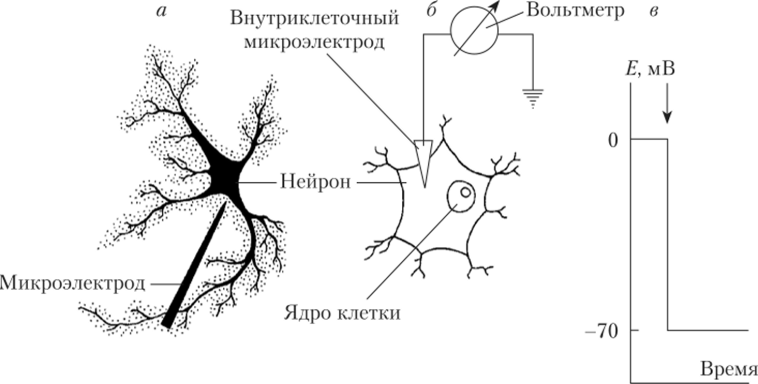
Рис. 8.1. Регистрация потенциала покоя нейрона:
а — рисунок с микрофотографии; б — схема регистрации потенциала покоя путем введения в нейрон кончика микроэлектрода; в — скачок мембранного потенциала в момент введения в нейрон кончика микроэлектрода (4).
В момент прокалывания мембраны нейрона регистрируется скачок потенциала вниз до уровня приблизительно -70 мВ. Это и есть мембранный потенциал, или ПП. Если электрод не двигать, а для нейрона создать правильные условия (состав окружающего раствора, температура), то ПП будет поддерживаться без всяких видимых изменений в течение нескольких часов. ПП найден во всех возбудимых клетках, и его величина колеблется от -30 до -100 мВ в зависимости от того, с какими клетками проводится опыт.
Потенциал покоя был открыт в середине XIX в. великим швейцарским физиологом Эмилем Дюбуа-РеймоноМу а его ученик Юлиус Бернштейн создал самую первую теорию, которая объясняла, почему внутри возбудимых клеток, находящихся в состоянии покоя, регистрируется явный избыток отрицательных зарядов. Исходя из данных, полученных целым рядом исследователей, Бернштейн частично установил, а частично предположил следующее:
- — в цитоплазме возбудимых клеток содержится гораздо больше ионов К+, чем в окружающей среде, а ионов Na+ и С1~ — гораздо меньше;
- — возбудимые клетки покрыты полупроницаемой мембраной, которая в состоянии покоя свободно пропускает ионы Na+, Са2+, С1_ и т. д.
Если это так, то часть ионов К+ должна выходить через клеточную мембрану в межклеточную среду, оставляя в клетке избыток парных К+ отрицательно заряженных частиц — анионов.
Поэтому на внутренней поверхности мембраны клетки создается отрицательный заряд (рис. 8.2).
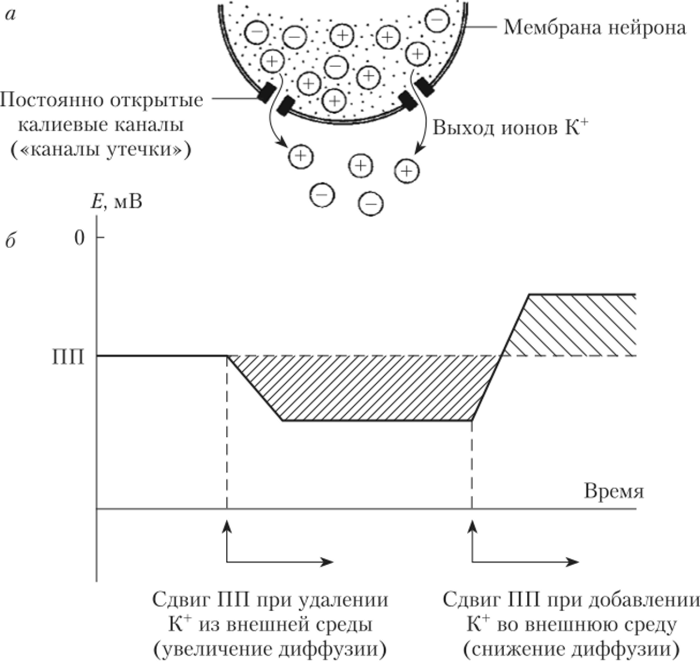
Рис. 8.2. Роль ионов К+ в возникновении и поддержании потенциала покоя (ПП):
а — возникновение ПП вследствие диффузии ионов К+ через постоянно открытые каналы утечки; б — изменение уровня ПП при изменении концентрации К* во внешней среде, Но избыток отрицательно заряженных частиц будет притягивать к себе ионы К+, препятствуя их выходу из клетки, а вышедшие в межклеточное пространство ионы К' также будут «отталкивать» положительно заряженные частицы, не давая выйти из клетки новым порциям К+. В итоге выход К+ продолжается лишь до тех пор, пока сила диффузии (концентрационного давления) и сила электрического ноля не станут равны. Точка равновесия и соответствует уровню потенциала покоя.
Ток ионов при достижении ПП не останавливается, так как имеются постоянно открытые каналы и К+ продолжает перемещаться сквозь мембрану, но число ионов, которые вошли в клетку и вышли из нее, теперь оказывается одинаковым. Такое состояние называется динамическим равновесием — равенством двух противоположно направленных процессов. Если один из процессов усилится или ослабнет, то точка равновесия сместится. Например, если, как это делал Бернштейн, искусственно увеличить концентрацию К+ в межклеточной среде, то это, естественно, будет мешать выходу из клетки новых порций положительно заряженных частиц (К+), и ПП станет менее отрицательным, смещаясь к нулю (см. рис. 8.2). Если же искусственно уравнять концентрацию К+ в цитоплазме и вне клетки, то ПП будет равен нулю. Так Бернштейн доказал, что ПП в возбудимой клетке определяется разницей концентрации К+ внутри этой клетки и во внеклеточной среде. Бернштейн предложил для количественной оценки ПП использовать уравнение, выведенное Вальтером Нернстом для искусственной системы, представляющей собой два раствора КС1 различной концентрации, разделенных искусственной мембраной, проницаемой только для К+ (рис. 8.3).
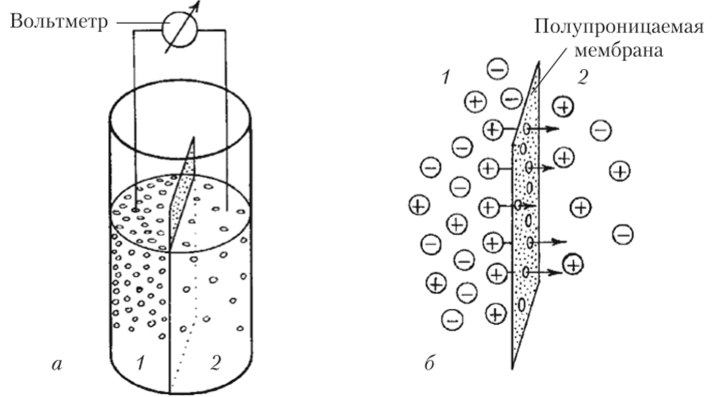
Рис. 83. Схема опыта Вальтера Нернста:
а — принципиальная схема установки с двумя сосудами (1 и 2), содержащими разные концентрации КС1 и разделенными мембраной, проницаемой только для К+;
6 — поток (показан стрелками) катионов (+) через мембрану из области с их большей концентрацией (1) в область с меньшей концентрацией (2), приводящий к возникновению на мембране потенциала В один сосуд он налил 10%-ный раствор КС1, а во второй — 1%-ный раствор этой соли. В обоих растворах произошла диссоциация КС1 на К+ и СП, но в сосуде 1 исходно было в 10 раз больше и катионов (К+), и анионов (С1″), чем в сосуде 2. Так как разделяющая растворы полупроницаемая мембрана хорошо пропускает катионы, то часть ионов калия (К+) перешла из сосуда 1, где концентрация КС1 исходно была выше, в сосуд 2, в котором концентрация КС1 была в 10 раз ниже. Поскольку ионы калия несут положительный заряд, то положительных зарядов в сосуде 2 будет больше, чем отрицательных. В сосуде 1 таким образом окажется некоторый избыток «брошенных» анионов хлора, потерявших свои катионы калия. Но отрицательно заряженные ионы С1″ будут притягивать назад часть положительно заряженных ионов К+ за счет взаимодействия электрических зарядов. Через какие-то мгновения потоки К+ из сосуда 1 в сосуд 2 и наоборот станут равными. Однако в сосуде 1 ионов К+ будет все равно меньше, чем ионов С1_ (ведь часть К* перешла в сосуд 2), а в сосуде 2 будет больше ионов К+, чем ионов С1_ (за счет пришедших из сосуда 1). Следовательно, раствор в сосуде 1 будет заряжен отрицательно по отношению к раствору в сосуде 2.
Между растворами установится нернстовский потенциал, который, согласно уравнению Нернста, равен.
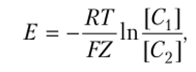
где Е — величина потенциала, возникающего между растворами, налитыми в сосуд 1 и сосуд 2; R — газовая постоянная; Т — температура; F — число Фарадея; Z — валентность; [С{ — концентрация ионов К+ в сосуде 1; [С2] концентрация ионов К+ в сосуде 2.
Бернштейн предложил использовать приведенное выше уравнение Нернста для определения мембранного потенциала, как только узнал, что именно К+ может проходить через мембрану покоящейся возбудимой клетки. Однако соотношение концентраций катионов калия внутри и снаружи клетки оказалось не 1: 10 (как в искусственной экспериментальной системе Нернста), а совсем другим. Так, в мышечных клетках калия в 49 раз больше, чем в среде, окружающей клетки. Зато в окружающей среде, т. е. вокруг возбудимых клеток, ионов натрия (Na+) приблизительно в 10 раз больше, чем внутри клеток. Однако, когда клетка находится в состоянии покоя, натрий не может проникать сквозь клеточную мембрану внутрь клетки и не участвует в создании 1111. За последние 70 лет многие ученые путем оригинальных экспериментов окончательно доказали, что отрицательный заряд покоящейся возбудимой клетки обусловлен тем, что часть ионов калия выходит через наружную мембрану во внешнюю среду до тех пор, пока не установится динамическое равновесие: сколько ионов калия вышло из клетки, столько же и войдет обратно. При этом на мембране устанавливается ПП, характерный для данного вида возбудимых клеток и определяемый тем, что в цитоплазме клетки остается избыток отрицательно заряженных ионов — анионов, главным образом, крупных белковых молекул, имеющих отрицательный заряд, «покинутых» катионами, т. е. ионами калия. Анионы пройти через мембрану не могут и остаются в клетке, определяя отрицательный заряд цитоплазмы.
Однако впоследствии было показано, что в мембране нейронов имеется небольшое количество постоянно открытых каналов для калия, по которым катионы натрия по градиенту концентрации (вокруг нейрона в 10 раз больше натрия, чем в цитоплазме) постоянно проникают в клетку, сдвигая 1111 к менее отрицательным значениям.
Входящий в клетку небольшой ток натрия (ток утечки), изменяя IIII, делает нейрон более возбудимым; чем этот ток значительнее, тем сильнее IIII сдвинут к тем пороговым величинам, за которыми следует переход нейрона в состояние возбуждения. Таким образом, возникает возможность поддерживать нейроны одних систем на высоком уровне возбудимости, а других — на пониженном. Как было выяснено, в межклеточной среде содержится в пять раз больше ионов хлора (С1″), чем в цитоплазме, однако в покое ток ионов хлора через наружную мембрану в клетку очень мал и составляет всего 4%, если сравнить с током ионов калия, но, строго говоря, для точного определения величины ГГП в уравнении Нернста надо подставить величины проницаемости для всех видов ионов, которые могут в покое проникать через мембрану клетки и менять ПП. Полученное уравнение носит имя Голдмана — Ходжкина — Катца:
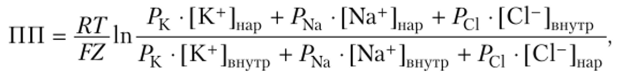
где Р — проницаемость мембраны для соответствующих ионов.
Механизм генерации ПП, концентрация ионов вне и внутри клеток, проницаемость наружной мембраны для ионов — все это исследовалось на различных объектах, однако основной вид клеток, на котором проводились опыты, — нейроны кальмаров (точнее, аксоны этих нейронов). В 1930;е гг. было обнаружено, что аксоны нейронов некоторых нервных узлов (звездчатого ганглия) этих головоногих имеют гигантский диаметр (до 1 мм), и с ним очень удобно экспериментировать.
Итак, ПП возбудимой клетки определяется несколькими факторами, главными из которых, безусловно, являются разница концентраций К+ внутри и снаружи клетки и свободный ток К+, идущий через мембрану. Определенную роль в генерации ПП играют слабые входящие токи Na+ и С1″, для которых мембрана клетки, находящейся в состоянии покоя, почти непроницаема. Изменяя любой из этих параметров, можно в той или иной степени менять ПП, тем самым изменяя способность клетки к возбуждению.