Транспозиции.
Генетика

Обычно хромосома Е. coli имеет несколько /5-элементов, например 8 копий /51, 5 копий /52 и т. д. Они перемещаются по хромосоме с частотой около 1 х Ю" 6 — 1 х КГ8 на клеточное деление. /5-элементы локализованы также в /'-факторе Е. coli два /53, один /52 и еще один элемент, обозначаемый уб. Именно по этим мигрирующим элементам и происходит рекомбинация, когда /" -фактор интегрирует с хромосомой… Читать ещё >
Транспозиции. Генетика (реферат, курсовая, диплом, контрольная)
Транспозиции представляют собой перемещение небольших участков генетического материала в пределах одной хромосомы или между разными хромосомами. Транспозиции происходят при участии особых подвижных или мигрирующих генетических элементов.
Впервые мигрирующие генетические элементы были описаны Б. Мак-Клинток в 1947 г. в связи с изучением хромосомных разрывов у кукурузы. Был обнаружен мигрирующий локус ?>$ (<диссоциатор), в котором предпочтительно происходят разрывы хромосом. Сам по себе ?>$ не вызывает разрывов. Они появляются в этом локусе, если в геноме присутствует другой мигрирующий элемент — Ас {активатор). Оба эти элемента могут теряться с частотой несколько процентов в мейотическом потомстве или менять свою локализацию при митотических делениях. При этом Ds перемещается только в присутствии Ас.
Внедрение Ds в непосредственной близости или внутрь гена С, контролирующего окраску алейрона семян, приводило к инактивации гена С, и гетерозиготные семена С/с/с (напомним, что эндосперм — триплоидная ткань) оказывались неокрашенными. В присутствии активатора (Ас) диссоциатор (Ds) начинал перемещаться — иногда покидал локус С. В результате этого появлялись окрашенные пятна алейрона на неокрашенных семенах (рис. 9.8).
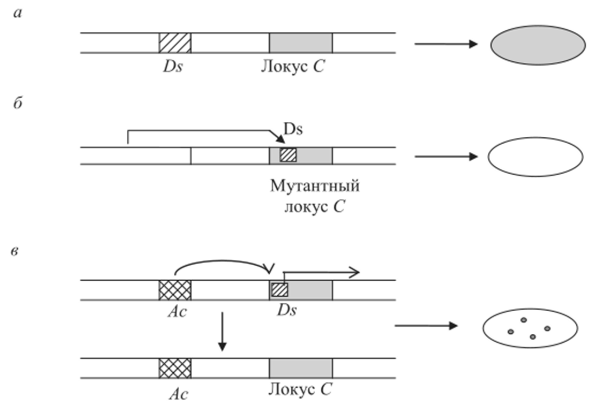
Рис. 9.8. Схема взаимодействия активатора (Ас) и диссоциатора ^), инактивировавшего локус С у кукурузы: аОз отдален от локуса С; бDs переместился в локус С и инактивировал его (проявляется как рецессивный аллель с); в-Ас стимулирует перемещения /)$, в результате чего покидает локус С, что выражается в реверсиях с->С в ходе митотических делений (появление окрашенных пятен на зернах кукурузы) Благодаря успехам генной инженерии в 1980;х гг. удалось выделить и исследовать Ас, Ds и некоторые другие мигрирующие элементы кукурузы (рис. 9.9). Оказалось, что Ds — это дефектный делегированный вариант Ас. Структура элемента Ас оказалась типичной для мигрирующих элементов, которые к этому времени были изучены прежде всего у бактерий, а также у дрозофилы и дрожжей БассИ. сегеч’шае.
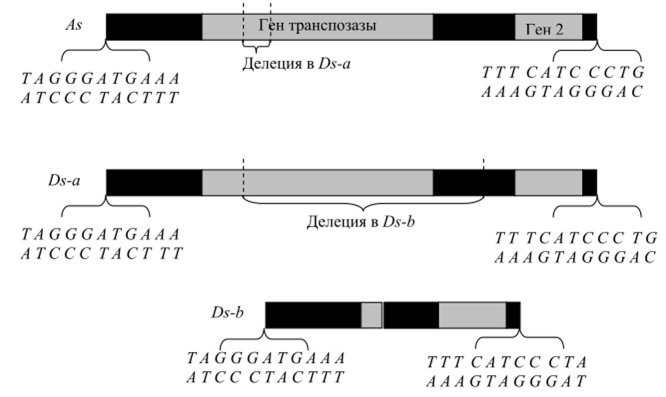
Рис. 9.9. Строение Л*- и Ду-элементов кукурузы. Показаны два гена (светлым), ответственные за транспозицию, и последовательности инвертированных несовершенных повторов на концах.
Начало изучению молекулярной структуры мигрирующих генетических элементов положило открытие в конце 1960;х гг. необычных мутантов по лактозному оперону Е. coli. У этих мутантов были инактивированы все три гена /дс-оперона. Частицы бактериофага А., трансдуцирующего /ос-оперон из таких мутантов, обладали необычайно высокой плавучей плотностью, что указывало на присутствие в lac-опероне мутантов лишней ДНК. Подобные мутации были найдены затем и в других генах Е. coli, фагах X и Р2. Общими для всех этих мутантов были инсерции (вставки) большей или меньшей длины. Эти вставляемые в разные участки генома Е. coli молекулы ДНК получили наименование /5-элементов (от англ, insertion sequences — вставные последовательности).
Размеры /5-элементов могут варьировать от 200 до 5 700 п.н. Все /5-элементы характеризуются следующими структурными особенностями:
- 1. На концах /5-элементы несут несовершенные, т. е. неидентичные инвертированные повторы нуклеотидной последовательности размерами от нескольких пар до нескольких десятков пар нуклеотидов.
- 2. Большинство /5-элементов содержит ген для фермента транспозазы, ответственной за их перемещение.
- 3. /5-элементы могут содержать по несколько сигналов начала и конца трансляции, а также сочетание нуклеотидов, сходное с сигналами терминации транскрипции.
4. В точке внедрения каждого /5-элемента, на его флангах всегда обнаруживается дупликация (в прямой ориентации) размером от 4 до 9 п.н. Эта дупликация не является частью /5-элемента, а представляет собой результат повторений сайта-мишени, в который внедряется элемент.
Обычно хромосома Е. coli имеет несколько /5-элементов, например 8 копий /51, 5 копий /52 и т. д. Они перемещаются по хромосоме с частотой около 1 х Ю" 6 — 1 х КГ8 на клеточное деление. /5-элементы локализованы также в /'-факторе Е. coli два /53, один /52 и еще один элемент, обозначаемый уб. Именно по этим мигрирующим элементам и происходит рекомбинация, когда /" -фактор интегрирует с хромосомой Е. coli, образуя ///•-штаммы. На это указывают результаты изучения ДНК /'-факторов, у которых участок ДНК бактериальной хромосомы, включенный в /'-фактор, оказывается отделенным от ДНК /'-фактора по обоим концам одной и той же последовательностью /5-элемента. /5-элементы /'-фактора и такие же последовательности, разбросанные по бактериальной хромосоме, создают условия для образования ////-доноров с различными началами и направлениями переноса бактериальной хромосомы.
Миграция /5-элементов, очевидно, связана с рекомбинацией, однако ее механизм отличен от классической гомологичной рекомбинации. На это указывает возможность транспозиции даже в клетках бактерий, несущих мутацию гес А, блокирующую общую гомологическую рекомбинацию у Е. coli.
В дальнейшем у бактерий были обнаружены более сложные мигрирующие элементы — транспозоны, которые отличаются от /5-элементов тем, что в них включены некоторые гены, не имеющие отношения к самому процессу транспозиции. Сходно устроены и транспозоны эукариот, например 7>1 — элемент Sacch. cerevisiae размером 5 700 п.н., вызывающий дупликации 5 п.н. в точках интеграции с ДНК хромосом (рис. 9.10). Подобное строение имеют и множественные диспергированные гены (МДГ) D. melanogaster, и ДНК-копии ретровирусов.
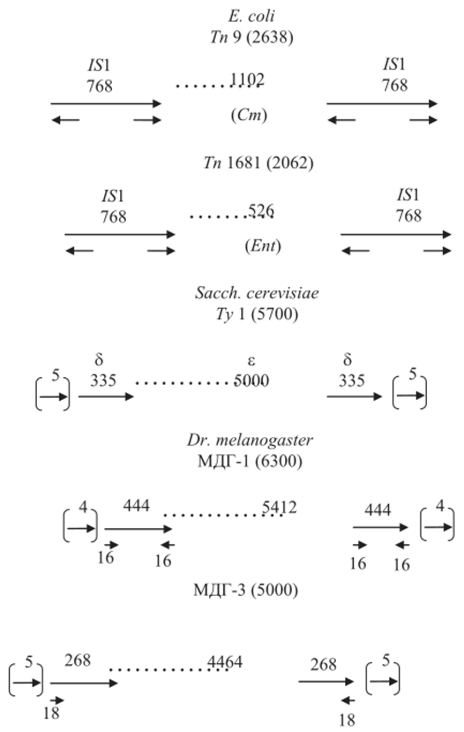
Рис. 9.10. Строение некоторых транспозонов Е. coli: Тп9 и Тп6Ъ Sacch. cerevisiae: 7>I; D. melanogaster: МДГ-1 и МДГ-3. Цифры — размер в числе пар нуклеотидов. Стрелки — прямые и инвертированные повторы. Стрелки в скобках — прямой повтор сайта-мишени. Cm — устойчивость к хлорамфениколу; Ent — продукция энтеротоксина; б, е — обозначения участков Ту.
Изучение нуклеотидной последовательности дуплицируемых сайтовмишеней на концах мигрирующих элементов показало, что они как правило неодинаковы как у различающихся элементов, так и у одного и того же элемента, локализованного в разных местах. Следовательно, мигрирующие элементы внедряются в те или иные точки генома независимо от их структуры. Правда, предпочтение во многих случаях отдается АТ-содержащим районам.
Некоторые мигрирующие элементы, покидая точку своей локализации, претерпевают внутрихромосомную гомологичную рекомбинацию по фланкирующим их длинным концевым повторам. В результате этого транспозон оставляет после себя одну копию своего концевого повтора, как это показано, например, для дрожжевого транслозона Ту (рис. 9.11), оставляющего после себя одну копию 5-повтора. По-видимому, сходно могут вести себя и МДГ О. melanogaster. Этим объясняется повторное появление некоторых мигрирующих элементов в оставленных ими сайтах — за счет гомологичной рекомбинации с оставшимся концевым элементом.
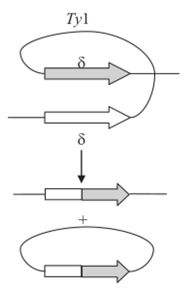
Рис. 9.11. Эксцизия (вырезание) дрожжевого транслозона Ту за счет рекомбинации между терминальными повторами элемента 8.
Для транспозонов эукариот рассматривают три механизма транспозиции:
- 1. Эксцизия предсуществующего транслозона с переносом на новое место — нерепликативная транспозиция.
- 2. Репликация ДНК транслозона с последующей транспозицией — репликативная транспозиция.
- 3. Обратная транскрипция РНК-копии транслозона и перемещение ДНК-копии на новое место — РНК-опосредованная транспозиция.
Обычно перемещения мигрирующих элементов не направлены и довольно редки. Они происходят с частотой 1×10-5 — 1×10″ 8, о чем можно судить по частоте ревертирования мутантов, возникших в результате внедрения этих элементов в непосредственной близости от генов или внутрь конкретных генов. В то же время можно создать такие экспериментальные ситуации, при которых частота транспозиции резко возрастает, а перемещения транспозонов перестают быть случайными. В линиях дрозофилы, длительное время (более 500 поколений) селектировавшихся на понижение половой активности самцов и общей жизнеспособности, отбирается неслучайный рисунок расположения МДГ в хромосомах. Об этом можно судить по гибридизации клонированных и радиоактивных копий МДГ с ДНК гигантских хромосом D. melanogaster непосредственно на цитологических препаратах слюнных желез личинок дрозофилы (in situ). Как только селекция в отрицательную сторону прекращалась, происходило быстрое (в течение нескольких поколений) перемещение копий МДГ, резко изменявшее картину их расположения на хромосомах. Это доказывает, что миграция транспозонов чутко отражает направление селекции и связана с адаптивной ценностью организма.
На основе этого появляются бактериальные эписом-факторы устойчивости, «собирающие» по несколько генов, обеспечивающих выживание бактерий в период массовой терапии антибиотиками.