Физиология шерстной продуктивности

В базальном слое происходят митотические процессы, которые обеспечивают появление новых поколений клеток взамен постоянно слущивающихся поверхностных слоев. Над зародышевыми клетками эпидермиса располагается слой шиповатых клеток. Основная масса клеток этого слоя имеет полигональную форму. Шиповатая клетка более дифференцирована, чем базальная. Поэтому в здоровой коже не отмечается митоза… Читать ещё >
Физиология шерстной продуктивности (реферат, курсовая, диплом, контрольная)
Кожа является важным и весьма многосторонним в функциональном отношении органом. Она защищает организм от механических повреждений и потери воды. В нормальном состоянии кожа непроницаема для микроорганизмов, а также ядовитых и вредных веществ. Кожный покров играет исключительно важную роль в регулировании температуры в организме, поддерживая ее на уровне, свойственном каждому виду животных. Кожа принимает участие в обмене веществ, через нее с потом удаляются из организма некоторые продукты распада. Кроме того, она выполняет функцию газообмена, является важным органом чувств, в котором сосредоточены осязательные, температурные и болевые нервные окончания, в ней формируется и растет шерсть.
Шерстная продуктивность овцы неразрывно связана со строением кожи и процессами, протекающими в ней. Кожа развивается из двух эмбриональных зачатков. Наружный ее слой — эпителиальный, или эпидермис, образуется из эктодермы, а соединительнотканная основа, или собственно кожа, — из мезенхимы, главным образом из так называемой кожной пластинки. Под дермой находится подкожный жировой слой, или подкожная клетчатка. Эпидермис представляет собой наружный слой кожи, состоящий из многослойного эпителия, в котором от базального, или росткового, слоя до самой поверхности происходит непрерывное движение клеток. Малодифференцированные зародышевые клетки в результате движения по направлению к поверхности кожи претерпевают целый ряд морфологических и химических изменений и, в конечном счете, превращаются в роговые клетки. В эпидермисе различают пять слоев, которые располагаются в такой последовательности: 1) ростковый (базальный, или зародышевый, слой); 2) слой шиповатых клеток; 3) слой зернистых клеток (первые три слоя составляют мальпигиев слой эпидермиса); 4) блестящий слой; 5) роговой слой. Необходимо отметить, что в эпидермисе кожи овец четко разграничить перечисленные слои, как правило, не удается. Самый внутренний слой, непосредственно прилегающий к соединительной ткани, — ростковый. Он представлен одним рядом кубических или цилиндрических клеток с круглыми и овальными ядрами. Остальные ряды клеток несколько уплощены.
В базальном слое происходят митотические процессы, которые обеспечивают появление новых поколений клеток взамен постоянно слущивающихся поверхностных слоев. Над зародышевыми клетками эпидермиса располагается слой шиповатых клеток. Основная масса клеток этого слоя имеет полигональную форму. Шиповатая клетка более дифференцирована, чем базальная. Поэтому в здоровой коже не отмечается митоза в шиповатом слое. За шиповатыми клетками находится 2−4 ряда зернистых клеток, содержащих включения кератогиалина. Блестящий слой, как и предыдущие, также состоит из нескольких рядов клеток, претерпевающих следующую стадию ороговения. Эти клетки заполнены особым блестящим веществом — элсидином, которое образуется из зерен кератогиалина. Элеидин представляет собой белок типа альбумина. В результате преломления света под микроскопом он выглядит светящимся. Элеидин является промежуточным продуктом между кератогиалином и кератином. Блестящий слой неоднороден по своему составу и постепенно сливается с нижними рядами рогового слоя. Это наиболее мощный слой эпидермиса, в котором насчитывается много десятков рядов окончательно ороговевших клеток. В нижней и средней зонах рогового слоя клетки прочно соединены между собой, а на поверхности они разъединяются в виде чешуек. Обычно роговые клетки состоят из кератиновой оболочки, но в отдельных случаях в центре клетки можно обнаружить остатки ядра. Роговое вещество клетки представлено белком кератином.
Как известно, ороговение, при котором клетки проходят стадию кератогиалинового и элеидинового превращения, называется мягкой кератинизацией. Процесс, протекающий при образовании кератинов стержня волоса или рогового слоя эпидермиса, носит название кератинизации, при которой из сульфгидрильных групп образуются дисульфидные. По типу мягкой кератинизации происходит ороговение клеток эпидермиса и сердцевинного слоя волоса, а по типу твердой — коркового слоя волоса, рогов, когтей, перьев и т. д. В связи с этим кератин коркового слоя классифицируют как твердый, в отличие от мягкого кератина ороговевшего эпидермиса. Твердый кератин монолитен, в то время как мягкий осыпается в виде чешуек (перхоть). Преобладающая часть серы, содержащейся в твердом кератине, присутствует в виде цистина, в то время как ороговевший эпидермис содержит равные количества цистина и метионина.
Таким образом, при ороговении эпидермиса главное событие — окисление сульфгидрильных групп в клетках мальпигиевого слоя в дисульфидные рогового слоя. Установлено, что весь мальпигиев слой содержит большое количество SH-групп, в то время как в роговом слое их мало или нет совсем. Необходимо отметить, что толщина эпидермиса непостоянна и зависит от ряда факторов. Имеются данные, свидетельствующие о породных особенностях. Кроме того, показана зависимость толщины эпидермиса от возраста, сезона и физиологического состояния организма.
В пилярном слое выделяют две зоны: подэпидермальную, находящуюся непосредственно под эпидермисом до основания сальных желез, а также промежуточную, расположенную от основания сальных желез до основания потовых желез. Большинство клеток соединительной ткани пилярного слоя составляют фибробласты, причем в подэпидермапьной зоне клеточных элементов вдвое больше, чем в промежуточной. Среди волокнистых структур подэпидермапьной зоны преобладают коллагеновые и эластиновые волокна. Кроме того, в ней хорошо развита сеть кровеносных капилляров.
Сетчатый, или ретикулярный, слой образован переплетением главным образом пучков коллагеновых волокон, обеспечивающих качество и прочность кожной ткани овчины. Кожа овец разных пород заметно различается по морфологии ретикулярного слоя. Так, например, у грубошерстных овец по сравнению с тонкорунными она характеризуется более сложной и плотной вязью пучков, большим их диаметром. Коллагеновые волокна прочнее, но менее эластичны. Каждое волокно имеет фибриллярное строение, диаметр отдельных фибрилл составляет в среднем 0,1 мкм. Основной белок коллагеновых волокон — коллаген. Толщина ретикулярного слоя непостоянна. У взрослых овец она зависит от пола, их конституциональных особенностей, физиологического состояния и действия сезонных факторов. Толщина ретикулярного слоя в значительной степени обусловливает толщину кожи в целом. Подкожная клетчатка — самый глубокий слой кожи. Она защищает нижележащие части от охлаждения и, кроме того, служит для запасов жира. Подкожная жировая клетчатка образована из жировых долек и окружающих их соединительнотканных прослоек, т. е. из крупных пучков коллагеновых и эластических волокон, а также клеток соединительной ткани. Благодаря рыхлому расположению указанных пучков волокон подкожная клетчатка обеспечивает подвижное прикрепление кожи к прилегающим частям.
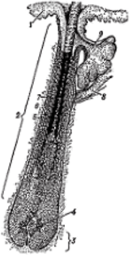
Рис. 3. Схема строения волоса (продольный разрез):
- 1 — эпидермис кожи;
- 2 — фолликул волоса;
- 3 — волосяная луковица;
- 4 — волосяной сосочек;
- 5 — кутикулярный слой.
- 6 — корковый слой,
- 7 — сердцевинный слой;
- 8 — мышца, поднимающая волос; 9 — сальная железа
Одной из важных функций кожи является продукция шерстных волокон. Корень волоса расположен в особом мешочке — волосяном фолликуле. Это микроскопических размеров специфические железы кожи, луковица которых служит секреторной частью, а наружное корневое (эпителиальное) влагалище — ее каналом. Строение волосяных фолликулов у овец (они отличаются постоянным ростом волос) такое же, как и у животных с чередованием периодов их роста и покоя. Волосяные фолликулы представляют собой удлиненные, цилиндрической формы образования, верхняя часть которых называется волосяной воронкой, а нижняя, закругленная, — волосяной луковицей, являющейся железистой частью фолликула, т. е. его матриксом, из которого растет волос. В волосяную луковицу вдается из кожи соединительнотканный сосочек, получивший название волосяного сосочка, несущий кровеносные сосуды, которые питают луковицу. Луковица представляет собой нижнюю, как бы расширенную часть корня. Напомним: в волосе различают стержень, т. е. часть, которая выступает над поверхностью кожи, и корень — часть волоса, располагающаяся в толще кожи между луковицей и стержнем. Корень волоса одет волосяной сумкой, которая построена из коллагеновых и эластических волокон, аморфного межуточного вещества и клеток. Она обильно снабжена кровеносными сосудами и нервами. Соединительнотканная сумка под луковицей образует сосочек волоса. От дна фолликулярной воронки до места открытия в фолликул выводного протока сальной железы эпителиальная стенка фолликула представлена наружным корневым (эпителиальным) влагалищем, которое в этой части фолликула имеет такое же строение, как и эпидермис, и, собственно, является его продолжением. По мере приближения к волосяной луковице наружное эпителиальное влагалище становится все тоньше и тоньше, наконец, редуцируется до одного ряда клеток, сливающихся с клетками луковицы. Клетки этого влагалища имеют светлую вакуолизированную цитоплазму вследствие наличия в ней большого количества гликогена. В настоящее время пока окончательно не решен вопрос о роли внешнего влагалища. По имеющимся данным, оно может выполнять трофическую или даже секреторную функцию. Значение внешнего корневого влагалища заключается в смазывании поверхности трения. Наружное влагалище волоса у ягнят и крыс, подобно внутреннему влагалищу волоса, представляет собой постоянно движущийся в сторону эпидермиса клеточный поток. Оно имеет свою зону пролиферации.
Ниже устья выводного протока сальной железы, между корнем волоса и наружным эпителиальным влагалищем, располагается внутреннее эпителиальное (корневое) влагалище, которое является производным волосяной луковицы и состоит из трех слоев: кутикулы, или кожицы, наружного слоя Генле и среднего слоя Гексли. На среднем и верхнем участках волоса все эти три слоя сливаются и эпителиальное влагалище здесь состоит исключительно из роговых клеток. Клетки кутикулы непосредственно примыкают к кутикуле корня волоса и поэтому имеют отпечаток его рисунка. Кутикула внутреннего эпителиального подобно кутикуле волоса построена из одного ряда черепицеподобно расположенных чешуек. На вертикальном срезе отчетливо видна взаимная сцепленность зубцов кутикул влагалища и волоса. Благодаря такому сцеплению стержень растущего волоса движется вместе с внутренним эпителиальным влагалищем. Слой Гекели состоит из 1−2 рядов клеток, которые содержат трихогиалин, окрашивающийся основными красками.
Наружный слой, или слой Генгле, состоит из одного ряда плоских клеток, заполненных также трихогиалином. По мере продвижения клеток этого слоя вверх трихогиалиновые зерна срастаются, образуя большие гомогенные глобулы. Слой Генгле непосредственно прилегает к внешнему эпителиальному влагалищу.
Процесс формирования фолликулов у овец условно можно разделить на пять стадий: формирование эпителиального зачатка, рост раннего фолликула, образование луковицы и сосочка, рост корня и влагалищ, выход стержня на поверхность кожи. Формирование в эпидермисе зачатков фолликулов происходит в период от 50- до 110-дневного внутриутробного развития плодов. Первоначальные зачатки волос у 50−70-дневных эмбрионов образуются в ростковом слое эпидермиса в виде скоплений эпидермальных клеток отдельными группами. Вначале зачатки имеют вид утолщений, которые со временем углубляются в соединительную ткань дермы, и, в конце концов, появляются расширения — будущие волосяные луковицы. В дальнейшем эти расширения обрастают соединительной тканью с кровеносными капиллярами (волосяные сосочки). Волосяной зачаток, в свою очередь, дает выпуклость, которая превращается в сальные железы. В зачатке волоса происходят сложные биохимические процессы, в результате которых и формируется сам волос.
Разнообразие шерстного покрова овец в значительной степени обусловлено различиями в строении фолликулов. Большинство исследователей разделяют волосяные фолликулы на первичные и вторичные. Основанием для такого деления в первую очередь служит последовательность появления фолликулов в процессе гистогенеза кожи плодов, их морфологические особенности, а также качество продуцируемых ими волокон. Первичные фолликулы образуются в ростковом слое эпидермального пласта. Они залегают глубоко в пилярном слое дермы. Каждый первичный фолликул имеет свою потовую железу, пучок гладких мышечных волокон, одну двухдольную или многодольную сальную железу. Волокна, продуцируемые первичными фолликулами, отличаются большим диаметром. У грубошерстных овец эти фолликулы образуют ость и мертвый волос, у тонкорунных новорожденных ягнят — песигу. Закладка первичных фолликулов происходит между 60−70 днями внутриутробного развития плода. Формирование их заканчивается в конце четвертого месяца развития плода. Вторичные фолликулы в отличие от первичных не имеют потовых желез и мускулов. Кроме того, сальные железы у них намного меньше их размеров. Эти фолликулы продуцируют пуховые волокна. Вторичные фолликулы образуются в том же ростковом слое эпидермального пласта, но намного позднее — в 70−110-дневном возрасте эмбрионов, и сам процесс закладки их не успевает заканчиваться к моменту рождения ягнят.
У овец грубошерстных пород первичные и вторичные фолликулы существенно различаются по форме, глубине залегания, строению луковиц и сосочков. Глубина залегания волосяных фолликулов в коже, а следовательно, толщина пилярного слоя связана с числом фолликулов на единицу площади кожи, диаметром и общей длиной их, а также состоянием самой кожи. Чем больше фолликулов на единице площади, тем они прямолинейнее, чем реже при той же длине и диаметре, тем больше их изогнутость, а отсюда и меньшая глубина залегания. Из длинных фолликулов, как правило, вырастает и длинная шерсть. Густота фолликулов на единицу площади кожи у тонкорунных овец в 2−3 раза большая, чем у грубошерстных. Число фолликулов на единице площади кожи с возрастом уменьшается, причем этот процесс интенсивнее происходит у баранов, чем у маток. Большие различия в густоте фолликулов имеются не только между породами, но и в пределах одной породы. От количества фолликулов на единицу площади кожи зависит густота шерстного покрова. Количество волокон на 1 см кожи у овец разных пород неодинаково: оно меньше у грубошерстных и больше у тонкорунных. По этому признаку большие колебания отмечаются у овец не только разных пород, но и внутри одной породы.
Волосяные фолликулы расположены не разрозненно, а пучками — волосяными группами. В каждой группе обычно есть один (реже 2 или 3) первичный фолликул и вокруг него несколько вторичных. У овец тонкорунных пород в волосяной группе количество вторичных фолликулов наибольшее, у грубошерстных — наименьшее. По числу первичных фолликулов волосяные группы разделяются на одинарные, двойные и тройные. В последних один из первичных фолликулов является центральным, или основным, а остальные два — боковыми (латеральными). Считают, что волосяные фолликулы закладываются исключительно в утробном периоде и часть их к моменту рождения ягненка уже продуцирует шерстные волокна, однако другие исследователи придерживаются мнения о том, что зачатки волосяных фолликулов формируются как в утробном, так и в постнатальном периоде.