Свет.
Физиология растений

Механизм действия света на рост до конца неясен; существует несколько гипотез. Первая гипотеза — прямое влияние света на активность фермента. Пигмент входит в состав фермента. Поглотив квант света, молекула пигмента переходит в возбужденное состояние, что вызывает изменение структуры связанной с ним белковой молекулы (апофермента) и, как следствие, активность фермента изменяется. Второй случай… Читать ещё >
Свет. Физиология растений (реферат, курсовая, диплом, контрольная)
Зависимость роста от света сложна, так как он выполняет две функции — субстратную и регуляторную. Субстратная роль света заключается в том, что он является источником энергии для фотосинтеза — процесса, создающего строительный материал для клеток и дыхательный субстрат. Однако прямой связи между скоростью роста и скоростью фотосинтеза нет: максимум роста наблюдается в течение вегетационного периода раньше, чем максимум фотосинтеза. У многих видов растений удлинение стеблей замедляется в дневные часы — в условиях, наиболее благоприятных для фотосинтеза (высокая освещенность), так как свет тормозит растяжение клеток. Эту закономерность впервые описал Ю. Сакс в 1872 г. (Германия) и назвал ее световым торможением роста. Однако более яркий свет, улучшая фотосинтез, может способствовать росту, несмотря на тормозящее действие света как регулятора. Тормозящее влияние света на рост является главной причиной суточной периодичности роста, о чем мы уже говорили. В то же время у растений, растущих в тени или в очень густых посевах, корневая система значительно слабее, чем у растений, получающих достаточное количество света.
Рост всех растений, в том числе и фотосинтезирующих, может продолжаться то или иное время в темноте за счет запасных веществ. Исключение составляет лишь небольшая группа растений, семена которых прорастают только под влиянием света. Однако в непрерывной темноте изменяется характер роста: тормозится деление клеток, и рост происходит за счет растяжения. В результате сильного растяжения клеток растения имеют длинные междоузлия. Темнота является сигналом, что растение, возможно, находится в почве и ему нужно выйти на поверхность. Поскольку в темноте хлорофилл не синтезируется, фотосинтез не идет, растение экономит строительный материал, и листовые пластинки недоразвиты. У растений плохо развита механическая и проводящая ткани, что связано с недостатком лигнина, в синтезе которого принимает участие фенилаланинаммиаклиаза (ФАЛ), активируемая светом. Такие растения называются этиолированными (рис. 7.26), а реакция на недостаток света — этиоляцией. Для этих растений характерны также слабый отрицательный геотропизм и сильный положительный фототропизм (см. параграф 7.4).
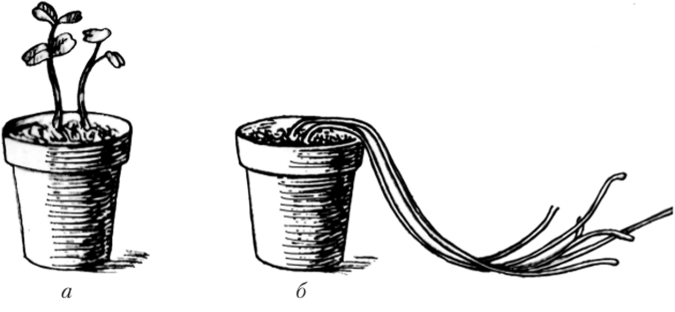
Рис. 7.26. Проростки белой горчицы, выращенные на свету (а).
и в темноте (б)1
Способность вытягивать в темноте стебли имеет большое значение в жизни растений: оказавшись глубоко под землей, проросток вытягивается до тех пор, пока не выйдет на поверхность почвы. На свету его рост[1]
в длину замедляется, и начинают формироваться настоящие листья, синтезируется хлорофилл. Происходит деэтиоляция.
Этиоляция не связана непосредственно ни с отсутствием хлорофилла, ни с недостатком питательных веществ. Так, при хранении в темноте клубней картофеля образуются этиолированные побеги, хотя питательных веществ хватает. С другой стороны, достаточно ежедневно освещать этиолированные растения в течение 5—10 мин даже довольно слабым светом, чтобы растяжение междоузлий начало тормозиться. Итак, этиоляция является приспособлением для быстрого выноса листьев на свет.
В слишком густых посевах злаков и некоторых других культур растения сильно затеняют друг друга, поэтому у них растягиваются нижние междоузлия и плохо развивается механическая ткань. Такие растения легко ложатся на землю — полегают. При высокой интенсивности света, в условиях активного фотосинтеза рост стебля замедляется, получаются невысокие растения с хорошо развитой механической тканью. Растущие в тени растения, например одуванчика лекарственного, обычно в несколько раз выше растущих на открытом месте.
У многих деревьев строение листа сильно зависит от освещения. Световые листья па периферии кроны значительно толще, чем теневые листья, расположенные внутри кроны, так как у них образуется больше рядов столбчатых клеток. У теневых листьев увеличено количество хлорофилла. Форма листьев также может зависеть от света. У колокольчика круглолистного при слабом освещении образуются круглые листья, а при сильном — узкие.
Влияние интенсивности света на рост сильно зависит от температуры: каждой температуре соответствует своя минимальная интенсивность света, причем разная, например для таких процессов, как заложение листьев и их линейный рост. С увеличением температуры возрастает и величина минимальной интенсивности света. Там, где комбинация этих факторов наиболее благоприятна, резко увеличивается рост.
Однако особенно заметна регуляторная роль света при освещении растений разным монохроматическим светом. Наиболее активно регулируют рост красные и синие лучи. Уже Г. А. Клебс (Германия, 1896) наблюдал, что синие и фиолетовые лучи стимулируют деление клеток, но тормозят их растяжение; красные лучи, наоборот, усиливают их растяжение. Зеленый свет, как и темнота, вызывает этиоляцию. Тормозящее влияние синего света на растяжение хорошо видно на примере растений высокогорных лугов. Эти растения всегда очень низкие, так как в горах на Землю попадает много сине-фиолетовых лучей. Даже если выращивать высоко в горах обычные для долин растения, они становятся похожими на высокогорные.
Спектральный состав света влияет и па другие особенности растений. Красный свет тормозит образование боковых корней, например у гороха, причем сильнее, чем синий и зеленый. У растений редиса только па синем свету образуется хороший корнеплод, а на красном — он практически не образуется. Причиной гетерофилии (от греч. heleros — другой, разный и pholio — лист), свойственной некоторым болотным и водным растениям, тоже считают влияние света разного качества, поскольку сине-фиолетовые лучи хуже проникают в толщу воды, чем красные. Например, у стрелолиста подводные листья имеют лентовидную форму, а надводные, образующие прикорневую розетку, — стреловидную. Если же эти растения освещать только красным или только зеленым светом, то у них образуются лишь лентовидные листья, а синим — нормальные листья.
Продолжительность освещения (длина дня) оказывает большое влияние на рост деревьев, причем и в этом случае растения разного географического происхождения реагируют тоже различно. У деревьев южных пород побеги на длинном дне долго растут, поэтому не успевают подготовиться к зиме и вымерзают. Деревья северных пород на длинном дне успевают хорошо подготовиться к зиме, замедлив рост.
Образование определенной пространственной структуры растения в результате действия света разной интенсивности, качества, продолжительности получило название фотоморфогеиеза.
Чтобы свет повлиял на рост, он должен быть поглощен. В конце 1940;х гг. Н. А. Бортвик и С. Хендрикс (США), изучая механизм действия света на прорастание семян салата-латука, открыли новый пигмент. Набухшие в темноте семена они помещали на слабый монохроматический свет различной длины волны. Оказалось, что быстрее всего семена прорастали при освещении их красным светом (X = 660 нм), значительно медленнее — при освещении синим и, наконец, практически не прорастали при освещении зелеными или дальними красными лучами (X = 730 нм). Поэтому возникло представление о том, что в семенах существует пигмент, который был выделен из растений и назван фитохромом.
Фитохром (ФХ) — пигмент из группы хромопротеидов. Его молекула состоит из двух белковых субъединиц: с молекулярной массой примерно 250 кДа и светопоглощающей хромофорной группой (от греч. скготаЬоя — цвет). Хромофорная группа содержит четыре пиррольных кольца, соединенных в цепь, — фитохромобилин (рис. 7.27).

Рис. 7.27. Структура фитохрома.
Фитохром существует в двух формах ФХ6б0 и ФХ730. ФХ660 — неактивная форма пигмента, голубого цвета, более устойчивая; ФХ730 — активная форма пигмента, зелено-желтого цвета, представляет собой восстановленную форму ФХ660. ФХбб0 поглощает красный свет с длиной волны 660 нм и переходит в ФХ730, а ФХ730, поглощая дальние красные лучи с длиной волны 730 нм, превращается в ФХб60. Упрощенную модель взаимопревращения различных форм пигмента и их действия можно представить так:
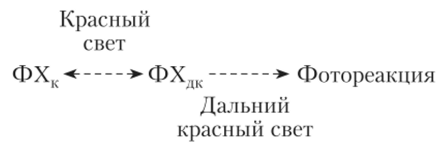
В темноте фитохром ФХ730 разрушается или переходит в неактивную форму ФХб60. Предполагают, что превращения фитохрома под действием света из неактивной формы в активную связаны с изменением положения тетрапиррола в пространстве и, соответственно, с изменением конформации связанного с ним белка, а также с фосфорилированием белка. Интенсивность красного света, необходимая для активации фитохрома, обычно очень низка — порядка 30 Дж/м2.
Неактивная форма фитохрома (ФХК) имеет длительный период жизни — около 100 ч, у активной формы этот период значительно короче — 30—60 мин. Фитохром — это растворимый белок, находящийся в цитозоле, при переходе в активную форму ФХЛК перемещается в ядро.
Оба фитохрома поглощают еще и синий свет: ФХ660 — с длиной волны 370 нм и ФХ730с длиной волны 400 нм.
Фитохром обнаружен не только в клетках всех органов растений, он есть у цианобактерий, грибов. Это говорит о том, что в процессе эволюции фитохромная система возникла давно. Больше всего его содержится в меристематических тканях.
Фитохром ФХ730 активирует гены, ферменты, увеличивает проницаемость мембран, участвует в регуляции таких процессов, как дифференцировка тканей и органов, рост стебля и гипокотиля, синтез антоциаиов, движения хлоропластов, прорастание семян, позеленение проростков (деэтиоляция), а также в фотопериодических реакциях, о которых будем говорить позднее (см. гл. 8). Фитохром и его превращения являются основой суточных ритмов фотосинтеза, дыхания, транспирации и других процессов.
У растений имеются регулируемые светом гены, активность которых подавлена в темноте специальными белками-репрессорами DET (от англ, de-etiolated) и СОР (от англ, constitutive photomorphogenesis). В темноте белок СОР накапливается в ядре и репрессирует (подавляет) транскрипцию генов, участвующих в фотоморфогенезе. При освещении свет поглощается фитохромом, передается на систему вторичных мессенджеров, включающую ионы кальция, кальмодулин, G-белки и др. Белок СОР выходит из ядра, репрессия светозависимых генов снимается, гены активируются, и продукты их экспрессии участвуют в фотоморфогенезе.
Реакции, управляемые фитохромом, зависят от концентрации и соотношения его активной (ФХ730) и неактивной (ФХ660) форм. Обычно реакция начинается, если 50% фитохрома представлено активной формой ФХ730. В полной темноте чувствительность растений к свету увеличивается, так как увеличивается содержание фитохрома. Как только растения переходят на свет, количество фитохрома уменьшается.
Действие фитохромов может изменяться в зависимости от уровня фитогормонов (цитокининов, брассиностероидов), а также при нападении патогенов.
Еще в конце XIX в. Ч. Дарвином (Англия, 1881) и Ю. Саксом (Германия, 1883) было установлено, что растения чувствуют синий свет. Однако рецепторы, воспринимающие синий свет, долго не могли выделить из клеток растений, отсюда они и получили свое название криптохромы (от греч. сгурШ — тайный, скрытый). В настоящее время известно, что в регуляции роста сипим светом участвуют два криптохрома и два фототропипа.
В 1993 г. А. Р. Кашмор и М. Ахмад доказали, что криптохромы являются хромопротеидами, хромофорная часть которых представлена флавинами (ФМН, ФАД) и птерином.
Птерин (метеиилтетрагидрофолат) функционирует как светособирающий хромофор. Интересно, что птерины являются пигментами глаз и крыльев насекомых. Криптохромы находятся в ядре и цитоплазме. Белки криптохромов родственны ДНК-фотолиазам — ферментам, восстанавливающим ДНК в присутствии света.
Кринтохромы поглощают синий (390—500 нм) и сильный зеленый свет. Предполагают, что они могут поглощать частично и ультрафиолетовые лучи (УФ-А с длиной волны 320—390 нм).
Под действием синего света кринтохромы фосфорилируются. При поглощении кванта синего света возбуждение передается на молекулу ФАД. Получив квант света, электрон флавина возбуждается, т. е. переходит на более высокий уровень, и редокс-потенциал резко изменяется. Возбужденный флавин — сильный восстановитель.
Криптохромы контролируют синтез антоцианов, каротиноидов, замедляют на свету рост гипокотиля или междоузлий, помогают фитохрому контролировать процесс деэтиоляции и определять продолжительность дня.
Фототропииы — рецепторы синего света, в отличие от криптохромов, содержат только флавин и соединены с мембранами. По химической природе они являются протеинкиназами, которые способны к автофосфорилированию в присутствии АТФ (см. подпараграф 1.6.2).
Все эти фоторецепторы влияют на активность транскрипционных факторов через цепь передачи сигнала, включающую изменение концентрации ионов кальция в цитозоле и активность протеинкиназ. Таким образом регулируется экспрессия генов.
Итак, основными фоторецепторами являются фитохромы, криптохромы и фототропииы. Фитохромы воспринимают красный и дальний красный свет, криптохромы и фототропииы — синий. Рецепторы синего и красного света взаимодействуют друг с другом.
Фотоморфогенез не связан непосредственно с фотосинтезом. Для устранения этиоляции и включения фотоморфогенеза достаточно ежедневного 5—10-минутного освещения.
Механизм действия света на рост до конца неясен; существует несколько гипотез. Первая гипотеза — прямое влияние света на активность фермента. Пигмент входит в состав фермента. Поглотив квант света, молекула пигмента переходит в возбужденное состояние, что вызывает изменение структуры связанной с ним белковой молекулы (апофермента) и, как следствие, активность фермента изменяется. Второй случай — косвенное влияние света на активность фермента. Молекула пигмента, поглотившая квант света, переходит в возбужденное состояние, взаимодействует с мембраной клетки и вызывает изменение ее структуры. В результате изменяется заряд мембраны, проницаемость и активность связанных с ней ферментов. Все это, в свою очередь, может быть причиной изменения метаболизма клетки и ее энергетических возможностей, изменения соотношения в клетке гормонов и ингибиторов роста, что в конечном счете приведет к изменению скорости процессов роста и дифференцировке.
Вторая гипотеза — непосредственное действие света на генетический аппарат. Пигменты, участвующие в регуляции роста растений светом, оказывают при возбуждении их светом непосредственное действие па гены, превращая неактивные в данный момент гены в активные, и наоборот. В результате синтезируются новые иРНК и белки, образование которых раньше было запрещено. Так, в ядре и хлоропласте фитохром влияет на гены, регулирующие синтез РуБисКО. Синий свет активирует в ядре гены, регулирующие синтез нитратредуктазы. Эта точка зрения с 1972 г. активно развивается в Германии крупным специалистом в области фотобиологии X. Мором.
- [1] См.: Максимов II. А. Краткий курс физиологии растений.