Влияние патогенов на физиологические функции

При нарушении одной из клеточных функций автоматически в патологический процесс будут включаться все новые функции клеток. Например, при увеличении проницаемости плазмалеммы нарушается ионный состав клетки, что, в свою очередь, изменяет структуру белковых глобул, следовательно, строение и функции митохондрий, ядра и вакуолей. Нарушение функций митохондрий, приводящее к недостатку АТФ, вызовет еще… Читать ещё >
Влияние патогенов на физиологические функции (реферат, курсовая, диплом, контрольная)
В растение патогены, как правило, проникают разными путями: через покровные ткани, состоящие из клеток, покрытых кутикулой (возбудители настоящих мучнистых рос, цветковые паразиты), большая часть — через устьица, чечевички, гидатоды, рыльце пестика (многие бактерии, возбудители ложных мучнистых рос), через микрои макротравмы (вирусы, многие грибы). Для дальнейшего распространения внутри растения патогену необходимо подавить защитные свойства уже не отдельной клетки, а ткани; изменить метаболизм растения в нужную для себя сторону; обеспечить свой ближний и дальний транспорт в растении.
Многие фитопатогенные грибы и бактерии обладают гидролитическими ферментами, разрушающими вещества клеточных стенок. При заражении растений грибы и бактерии образуют большой набор этих ферментов с высокими активностями. Среди ферментов имеются пектиназы, целлюлазы, гемицеллюлазы, кутиназы, разрушающие клеточные стенки растения, а также вызывающие мацерацию тканей. Мацерация (от лат. maceratio — размягчение) — это искусственное или естественное разъединение клеток из-за разрушения срединных пластинок. Мацерация способствует быстрому проникновению патогена в растительные ткани, а продукты гидролиза клеточных стенок служат для него питательными веществами.
Попав на благоприятный для питания субстрат, патоген начинает размножаться, при этом он может находиться внутри клеток растения, в межклетниках, на поверхности пораженной ткани. Распространение патогенов может быть локальным или охватывающим значительные участки ткани, располагающиеся по всему растению.
Развитие заболевания многоклеточного организма начинается с клетки или группы клеток, а в клетках — с изменения свойств мембран. Сначала патоген или его метаболиты индуцируют окислительный взрыв (см. параграф 4.4). Постоянное образование активных форм кислорода является нормой и происходит в хлоропластах, митохондриях и пероксисомах, а также на клеточной стенке и плазмалемме; однако в ответ на стресс их количество резко возрастает.
Японские1 и российские[1][2] исследователи установили, что ускоренное образование активных форм кислорода, например в тканях картофеля, в ответ на заражение оомицетом Phytophtora infestans, наблюдается уже через 1 ч после контакта патогена с растением, а максимум их образования совпадает с началом гибели клеток, т. е. через 4—6 ч после заражения. Предполагают, что образование 02_ происходит на плазмалемме клеток растения-хозяина.
Активные формы кислорода вызывают перекисное окисление фосфолипидов мембран. Перекисное окисление липидов — сложный процесс, состоящий из нескольких этапов. Прежде всего, в тканях активируется липоксигеназа — фермент, катализирующий окисление ненасыщенной связи в молекуле жирной кислоты в насыщенную. Субстратом для этого фермента служат полиненасыщенные жирные кислоты, входящие в состав фосфолипидов мембран:
Полиненасьпценный жир + 02 —> Перекись ненасыщенного жира В результате в мембранах уменьшается содержание ненасыщенных жирных кислот, накапливаются продукты переокислсния липидов, в том числе малоновый диальдегид (МДА). Увеличение содержания насыщенных жирных кислот в мембране приводит к уменьшению ее текучести, что, в свою очередь, вызывает нарушение всех функций мембран. Хотя перекисное окисление мембранных фосфолипидов происходит и в нормальной клетке, его усиление является одним из наиболее распространенных механизмов повреждения структуры мембран под действием патогенов и других стрессоров.
Принято рассматривать три основных первичных механизма повреждения биомембран в результате перекисного окисления липидов. Во-первых, в гидрофобном слое мембраны появляются гидрофильные (гидроперекисные) группы, поэтому гидрофобность липидного бислоя нарушается, при этом резко увеличивается его проницаемость для ионов и активируется транспортная АТФаза. Это вызывает увеличение потоков ионов калия и протонов через мембраны, активацию калиевых и анионных насосов для электролитов, приводит к вымыванию ионов из клетки, изменению скорости и направленности транспорта метаболитов между отдельными органеллами, нарушению гомеостаза, снижению pH в клетке и его повышению во внеклеточном пространстве и в конце концов к гибели клетки. Во-вторых, перекисные радикалы окисляют аминокислотные остатки мембранных белков, особенно содержащие БН-группы, которых много в мембранных белках. При окислении БН-групп между ними могут образовываться серные мостики, что приводит к агрегации молекул белков в мембране. В-третьих, если эти БН-группы находятся в активном центре фермента, то фермент теряет свою активность. Взаимодействие АФК с белками, нуклеиновыми кислотами приводит к мутагенезу.
Мы уже говорили (см. параграф 4.6), что одной из главных мишеней для АФК является фотосинтетический аппарат. При действии АФК нарушаются структура и функции хлоропластов, происходит инактивация фотосистемы II, снижается активность РуБисКО и других ферментов, уменьшается концентрация хлорофиллов и каротиноидов. Повреждение фотосинтетического аппарата вызывает еще большее накопление АФК и, как следствие, повреждение мембран, цитоскелета, нарушение водного обмена.
Итак, перекисное окисление мембранных фосфолипидов является одним из наиболее распространенных механизмов разрушения мембранных структур.
Окислительный взрыв, вызванный патогеном, нарушает баланс между образованием АФК в норме и функционированием антиоксидантной системы растения.
С образованием активных форм кислорода, окислением липидов и нарушением окислительного фосфорилирования связано усиление сверхслабого свечения живых клеток. Сверхслабое свечение (ССС) — это излучение света в видимой области спектра живыми клетками. В обычных условиях оно настолько слабо, что не видно невооруженным глазом, отсюда и получило свое название. Сверхслабое свечение, а следовательно, и АФК, распространены в живой природе повсеместно. Однако любое действие на клетку патогенов и других стрессоров самой разной природы вызывает усиленное его образование, которое может продолжаться от нескольких секунд до нескольких часов. По характеру ССС можно судить об устойчивости растения. ССС — один из признаков окислительного взрыва.
Болезни связаны не только с образованием АФК, но и с действием мощных токсинов, которые патоген секретирует по мере размножения в клетках хозяина (см. параграф 10.1). По механизму действия токсины делят на две группы: токсины — ингибиторы ферментов и токсины, повреждающие мембраны.
Клеточные мембраны являются первой мишенью не только для действия АФК, но и для токсинов. Токсины ряда патогенов способны деполяризовать плазмалемму растения, они влияют на транспорт ионов через мембраны, индуцируют потерю клеткой важных метаболитов, некоторые токсины сами являются источником АФК. Под влиянием патогенных микроорганизмов из листьев, например хлопчатника, банана, выделяются в большем количестве калий, натрий, магний. Нарушаются функции мембранных рецепторов.
У растений токсины вызывают набухание пластид и митохондрий, уменьшение размера ядер. Набухание хлоропластов приводит к разобщению фотофосфорилирования и транспорта электронов и, как следствие, к снижению синтеза АТФ. Изменяются белки митохондрий: снижается их способность связывать липиды, в них обнаруживается большее количество БН-групи, чем в мембранных белках здоровых растений. Грибные токсины могут вызвать увеличение концентрации ненасыщенных липидов в мембранах корней и побегов на 62—65%, что приводит к торможению роста.
Токсины вирусов, так же как дефицит ионов Бе, Си и других элементов минерального питания, вызывают разрушение хлорофилла. Из-за низкого содержания хлорофилла изменяется окраска листьев: они становятся светло-зелеными, желтыми (хлороз) или на них появляются отдельные пожелтевшие участки (мозаика). Нарушается синтез белков и углеводов.
Под действием токсинов патогена не только увеличивается интенсивность дыхания, но и изменяется его химизм — активируется пентозофосфатный окислительный путь.
Нарушение функций митохондрий связано с изменением состава и соотношения липидов и белков мембран. При уменьшении числа двойных связей в липидах митохондрий активируется фосфолипаза А, которая катализирует гидролиз фосфолипидов, т. е. разрыв эфирных связей между жирной кислотой и глицерином. Этот процесс происходит и при старении хлоропластов.
Происходящее под действием фосфолипазы А изменение липидов вызывает набухание митохондрий, что становится причиной разобщения окисления и фосфорилирования, в митохондриях почти не синтезируется АТФ, что приводит к большему выделению тепла при дыхании. В результате температура листьев, пораженных, например, грибом-возбудителем корневой гнили, может повыситься на 3—5°С по сравнению с температурой здоровых листьев. Уменьшение степени ненасыщенности жирных кислот (ожирение мембран), увеличение количества стеринов лишает мембраны пластичности и способствует более легкой повреждаемости.
Некоторые патогенные бактерии и грибы могут индуцировать образование цианидов, которые вызывают увеличение активности альтернативной оксидазы, а следовательно, возрастание интенсивности альтернативного (цианидустойчивого) дыхания.
К токсинам относят и гидролитические ферменты. Грибы выделяют экзополигалактуропазы, вызывающие распад (деполимеризацию) полисахаридов клеточных стенок растения с образованием олигосахаридов.
Олигосахариды (от греч. oligos — малый) — полисахариды, содержащие в молекуле небольшое число остатков простых сахаров. Они образуются в результате деполимеризации полисахаридов клеточной стенки. Расщепление вызывают ферменты грибов и бактерий, а также самого растения (см. параграф 10.3). Некоторые олигосахариды обладают физиологической активностью, их называют олигосахарипами. В состав олигосахаринов могут входить ксилоза, рамноза, галактоза и остатки уроновых кислот. К олигосахаринам относят и короткие фрагменты хитина грибов, образующиеся под действием хитиназ. Например, когда патогенный гриб Fusarium solani атакует растение гороха, споры гриба секретируют очень небольшое количество кутиназы, которая вызывает частичный гидролиз кутикулы, покрывающей лист. Образовавшиеся мономеры кутина стимулируют усиленное образование кутиназы спорами гриба. В результате кутикула растворяется, и прорастающие споры могут проникнуть внутрь.
Вещества, вызывающие защитные реакции растения на контакт с патогеном, называют элиситорами (от англ, elicit — вызывать), или индукторами (от лат. inductor — побудитель). Элиситором может быть молекула белковой, углеводной, углеводно-белковой или липидной природы, не только свидетельствующая о присутствии патогена в растении, но и управляющая работой генов защиты. Элиситоры находятся на поверхности патогена или входят в состав его выделений, они действуют в очень малых концентрациях (КН М) и включают защитные реакции; некоторые элиситоры могут деполяризовать мембрану.
Одним из первых хорошо охарактеризованных элиситоров является гепта-[3-глюкозид (т.е. состоящий из семи остатков глюкозы), выделяющийся из клеточной стенки оомицета Phytophtora megasperma, поражающего сою (рис. 10.1). Он активирует ряд генов сои, в том числе принимающих участие в синтезе фитоалексинов (см. параграф 10.3). Хорошо изученным элиситором является липогликопротеидный комплекс. Он был выделен из оомицета-паразита Phytophtora infestans.
Большинство известных элиситоров являются неспецифическими, т. е. они вызывают защитные реакции у всех сортов растений и присутствуют у грибов всех рас. Растение узнает элиситоры и таким способом получает информацию о нападении патогена.
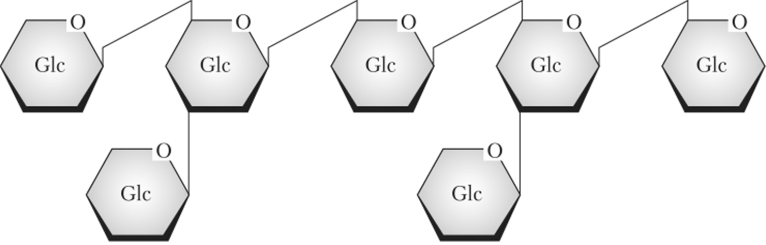
Рис. 10.1. Элиситор гепта-рглюкозид1.
Кроме того, в клетках самой ткани растения под влиянием инфекции усиливаются гидролитические процессы. К гидролазам относятся фосфатазы, катализирующие гидролиз сложных эфиров фосфорной кислоты и широко распространенные в микроорганизмах и растениях. В результате некротрофы превращают клеточное содержимое в удобную для питания патогена форму.
Также может происходить лигнификация или суберинизация клеточных стенок, что приводит к нарушению межклеточных связей, нарушению ближнего транспорта и метаболизма и в конце концов к отмиранию тканей — некрозообразованию. Некрозообразование — универсальная, неспецифическая реакция на заражение или повреждение. Одна из первопричин быстрой некротизации тканей растения — образование АФК на плазмалемме и связанное с ним переокисление липидов мембран.
Патоген оказывает сильное влияние на общий характер обмена веществ восприимчивого растения, направляя весь метаболизм в нужную для него сторону.
Вызванные токсинами бактерий изменения клеточных стенок, образование тилл, закупорка сосудов нарушают дальний транспорт воды по растению, что приводит к потере тургора и увяданию молодых побегов и листьев (рис. 10.2). Увядание листьев — типичный симптом заболевания растения, вызванный токсином. В то же время транспирационный ток может помогать распространению спор грибов.
В инфицированных тканях изменяется вторичный метаболизм. Например, происходит накопление фенолову прежде всего за счет их синтеза de novo. В процессе биосинтеза фенолов участвуют фенилаланинаммиаклиаза (ФАЛ), пероксидаза, нолифенолоксидаза, активность которых при повреждении клетки возрастает. Увеличение содержания фенолов может быть результатом также гидролиза глюкозидов, агликоном которых могут быть фенольные соединения. Кроме того, иод влиянием патогена активируется пентозофосфатный окислительный путь дыхания, промежуточным продуктом которого является эритрозо-4-фосфат — предшественник ароматического кольца, нужного для синтеза фенолов.
1 См.: Молекулярная биология клетки / Б. Альберте [и др. J: в 3 т. М.: Мир, 1994.
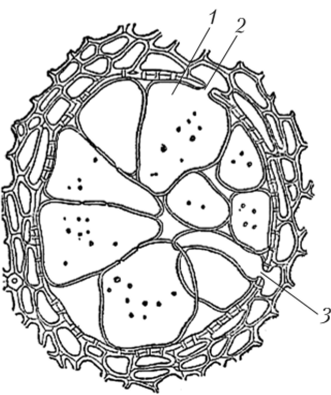
Рис. 10.2. Поперечный разрез сосуда, закупоренного тиллами1:
1 — поры на горизонтальных стенках тилл; 2,3 — отверстия пор, через которые в полость сосуда проникли выросты паренхимных клеток, ставших тиллами Фенолы быстро окисляются полифенолоксидазой и пероксидазой, и образуются хипопы. Хиноны подавляют активность дегидрогеназ, в результате процесс окисления фенолов становится необратимым и сдвинутым в сторону образования хинонов, которые повреждают мембраны лизосом. Гидролитические ферменты последних попадают в цитозоль и вызывают самопереваривание клетки (автолиз). Хиноны легко взаимодействуют с некоторыми группами белков (NH2-, ОН-, SH-) и тем самым могут инактивировать ферменты, а также повреждать белки мембран. Хиноны разобщают окисление и фосфорилирование.
Известно, что патогены синтезируют и выделяют в окружающую среду избыточное количество гормонов, что приводит к нарушению соотношения гормонов в растении, т. е. к нарушению гормонального статуса растения (см. параграф 7.2). Например, при контакте с Agrobacterium tumefaciens в геном нормальных клеток растения внедряются гены бактерии, в результате сами растительные клетки начинают интенсивно синтезировать ИУК и зеатин. Многие грибы вырабатывают ауксины, цитокинины, гиббереллины; бактерия Pseudomonas savastanoi синтезирует ауксин и цитокинин. Кроме того, под действием патогена в самом растении индуцируется синтез таких гормонов, как салициловая и жасминовая кислоты, системны, этилен.
Некоторые токсины сами функционируют как гормоны. Принадлежность гиббереллинов к фитогормонам давно установлена, а вопрос о гормональной природе фузикокцина до сих пор остается открытым. Фузикокцин, прежде всего, влияет на транспорт веществ через мембраны, увеличивает поступление воды в замыкающие клетки, что приводит к открыванию устьиц. Гриб Fusarium amygdali вырабатывает фузикокцин в огромных количествах. В результате увеличивается проницаемость мембран, акти;
1 См.: Курсанов Л. И. Курс ботаники: в 2 т. М.: УЧПЕДГИЗ, 1940.
вируется катион-стимулируемая АТФаза, что приводит к раку, например у миндаля и персика.
Гормоны патогена изменяют направление транспорта поглощенных корнями питательных веществ и оттока продуктов фотосинтеза, в результате происходит нарушение роста.
Избыточные концентрации гормонов, возникающие благодаря патогенам, могут вызвать изменение формы отдельных органов или всего растения. Деформация проявляется в виде скручивания, морщинистости, курчавости или нитевидности листьев, махровости цветков, уродливости плодов, укорачивании междоузлий. Деформация побегов обычно сопровождается повышенной ветвистостью, нарушением апикального доминирования — образованием множества тонких мелких побегов (ведьмины метлы). Некоторые патогены вызывают у растений образование галлов, вздутий, наростов, опухолей. Гормоны могут вызывать гипертрофию — увеличение размеров и изменение формы органов, клеток, гиперплазию — увеличение числа клеток, гипоплазию — уменьшение числа клеток и их размера, некроз, мацерацию.
Между токсинами и гормонами нет строгого разделения: токсичное или гормональное действие вещества зависит от его концентрации, вида растения, его возраста, органа, который подвергается действию.
Паразитам из числа высших растений обычно не придают большого значения, однако при сильном поражении они могут извлекать из проводящих тканей растения-хозяина такое количество воды и питательных веществ, что недостаток их становится ощутимым.
Физиологические и анатомо-морфологические изменения влияют на продуктивность растений и качество урожая. Снижение урожая и ухудшение его качества происходит также из-за стерильности цветков, опадания цветков и завязей, образования бессемянных плодов, связанного с нарушением соотношения гормонов.
Итак, под влиянием патогенов изменяются скорости дыхания, фотосинтеза и первичного метаболизма, нарушается структура и проницаемость мембран, углеводный, белковый, водный обмены.
При нарушении одной из клеточных функций автоматически в патологический процесс будут включаться все новые функции клеток. Например, при увеличении проницаемости плазмалеммы нарушается ионный состав клетки, что, в свою очередь, изменяет структуру белковых глобул, следовательно, строение и функции митохондрий, ядра и вакуолей. Нарушение функций митохондрий, приводящее к недостатку АТФ, вызовет еще большее увеличение проницаемости мембран, замедление энергозависимых процессов. Если избирательно подавить синтез белков, то это приведет к снижению синтеза РНК, ДНК из-за недостатка ферментов и структурных белков и к уменьшению активности митохондрий, а дальше — к изменению проницаемости мембран и набуханию клетки и т. д.
В зависимости от интенсивности, продолжительности и характера повреждения клетка может или адаптироваться к повреждающему фактору, реиарировать повреждение и восстановить свои функции, или резко снизить урожай и даже погибнуть.
Контрольные вопросы
- 1. Что такое окислительный взрыв?
- 2. Какие изменения в клетке вызывают активные формы кислорода?
- 3. Что такое перекисное окисление мембранных липидов?
- 4. Какие процессы являются следствием перекисного окисления липидов мембран?
- 5. Как изменяется дыхание клетки под влиянием инфекции?
- 6. Что такое сверхслабое свечение?
- 7. Как изменяется фотосинтез при действии на растение патогена?
- 8. Что происходит с клеточной стенкой под действием токсинов?
- 9. Какие гидролазы образует патоген?
- 10. Как изменяется водный обмен больного растения?
- 11. Как изменяется вторичный метаболизм в условиях атаки патогена?
- 12. Как изменяется гормональный статус пораженного патогеном растения?
- 13. Как изменяется рост растения и его органов при воздействии биотоксинов?
- 14. Как влияет инфекция на урожай и его качество?
- [1] См.: Doke N. Involvement of superoxide anion generation in hvpersensitive response ofpotato tuber tissues to infection with an incompatible race of Phytophthora infestans // Physiol. Plant Pathol. 1983. V. 23. P. 359−367.
- [2] См.: Аверьянов А. А. Генерация листьями риса радикала 02_ в связи с устойчивостьюк пирикуляриозу / А. А. Аверьянов [и др. J // Физиология растений. 1987. Т. 34. С. 373—379.