Структура ДНК.
Физиология растений в 2 т. Том 1

При исследовании химического состава ДНК, выделенных из разных организмов, органов и тканей, Э. Чаргафф (США) в 1950 г. установил, что в любой молекуле ДНК количество молей аденина равно количеству молей тимина, а количество молей цитозина точно равно содержанию гуанина; сумма пуриновых оснований в ДНК любого происхождения равна сумме пиримидиновых оснований. Эта закономерность получила название… Читать ещё >
Структура ДНК. Физиология растений в 2 т. Том 1 (реферат, курсовая, диплом, контрольная)
Нуклеиновые кислоты были впервые выделены из ядер спермы лосося и гноя швейцарским биохимиком Фридрихом Мишером в 1868—1872 гг. и названы им нуклеином (от греч. nucleus — ядро). Одновременно Ф. Мишер высказал идею, что нуклеиновые кислоты могут иметь отношение к передаче генетической информации. Эта идея получила блестящее подтверждение спустя 75 лет в опытах М. Маккарти (США). Маккарти продемонстрировал, что экстрагированная из болезнетворного штамма пневмококка ДНК несет генетическую информацию вирулетности, которая и передается здоровым клеткам, вызывая заболевание.
Нуклеиновые кислоты относятся к классу биологических полимеров. Их мономерами являются нуклеотиды, которые состоят из азотистого основания, пятиуглеродного сахара (пентозы) и остатка фосфорной кислоты (рис. 2.2). Нуклеотид без фосфатной группы называется нуклеозидом.
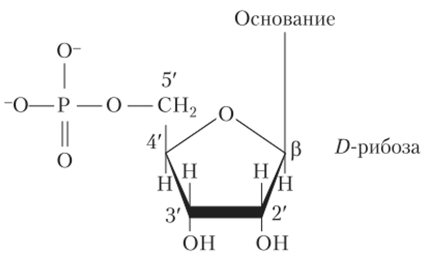
Рис. 2.2. Обобщенная структура нуклеотида.
Углеводные остатки представлены в ДНК 2−0-дезоксирибозой. Азотистое основание связано iV-гликозидной связью с 1 '-углеродом пентозы, а фосфатная группа — сложноэфирной связью с 5'-углеродом пентозы. В состав ДНК входят четыре азотистых основания: два пуриновых — аденин и гуанин (А, G) и два пиримидиновых — тимин и цитозин (Т, С) (рис. 2.3).
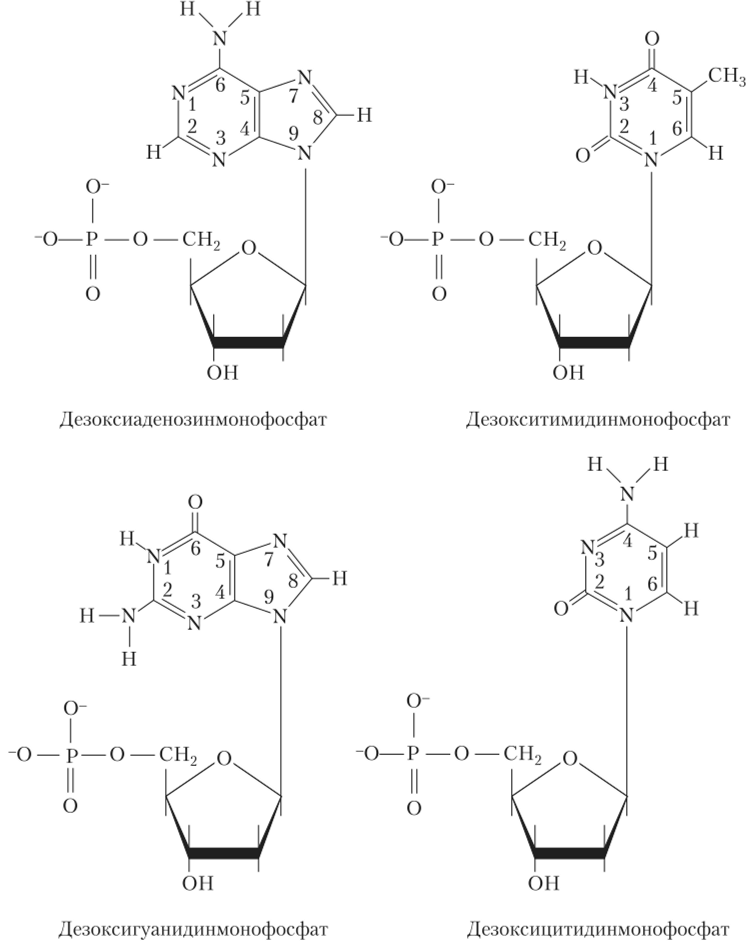
Рис. 2.3. Дезоксирибонуклеотиды:
цифрами обозначены положения атомов в гетероциклических кольцах пуринов (аденин и гуанин) и пиримидинов (тимин и цитозин) и углеродных атомов в дезоксирибозе
Нуклеотиды соединяются с помощью фосфодиэфирных связей, при этом фосфатная группа 5'-углеродного атома одного нуклеотида связана с З'-ОН-группой пентозы соседнего нуклеотида, в результате образуется цепочка (рис. 2.4). Концы цепочек обозначаются как 5'-конец и З'-конец. На 5'-конце цепи ДНК находится фосфатная группа, а на З'-конце — ОН-группа. Межнуклеотидные связи в ДНК и РНК расщепляются большим количеством разных пуклеаз. ДНК заряжена отрицательно за счет отрицательно заряженных остатков фосфорной кислоты.
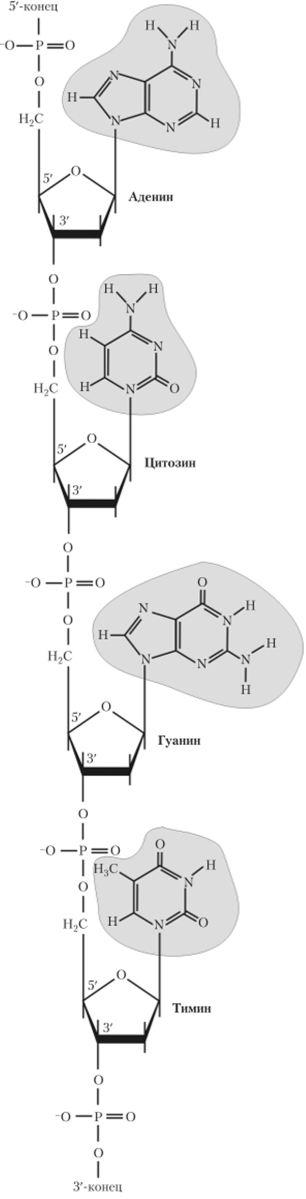
Рис. 2.4. Образование цепи из нуклеотидов.
В состав ДНК входят еще минорные основания (5-метилцитозин и 7-метилгуанин). Они обеспечивают правильное считывание генетической информации и позволяют нуклеазам отличать свою нуклеиновую кислоту от чужой.
При исследовании химического состава ДНК, выделенных из разных организмов, органов и тканей, Э. Чаргафф (США) в 1950 г. установил, что в любой молекуле ДНК количество молей аденина равно количеству молей тимина, а количество молей цитозина точно равно содержанию гуанина; сумма пуриновых оснований в ДНК любого происхождения равна сумме пиримидиновых оснований. Эта закономерность получила название правила Чаргаффа.
В 1953 г. американец Джеймс Уотсон и англичанин Фрэнсис Крик постулировали (установили) вторичную структуру ДНК, основываясь на данных рентгеноструктурного анализа кристаллов ДНК и на правиле Э. Чаргаффа. В соответствии с их моделью молекула ДНК представляет собой спираль, состоящую из двух полимерных цепей, закрученных друг относительно друга и вокруг общей оси (рис. 2.5), поэтому эта структура получила название двойной спирали. Напротив З'-копца одной цепочки в двухцепочечной молекуле ДНК располагается 5'-конец другой цепочки, т. е. эти цепи антипараллельны, направлены в противоположные стороны.
Гидрофильные остовы цепей, состоящие из чередующихся остатков дезоксирибозы и отрицательно заряженных фосфатных групп, расположены на внешней стороне двойной спирали, а гидрофобные основания — внутри спирали, стопками, перпендикулярно длинной оси (см. рис. 2.4). Отсюда следует, что основания одной цепи должны быть очень сильно сближены с основаниями другой.
Азотистые основания обеих цепочек располагаются друг против друга так, что против большего (имеющего двойной гетероцикл) пуринового основания одной цепи всегда располагается меньшее по размеру (имеющее один гетероцикл) пиримидиновое основание другой цепи, т. е. аденин одной цепочки располагается против тимина другой, а гуанин — против цитозина. Таким образом, полинуклеотидные цепочки комплементарны. Никакое другое сочетание оснований невозможно.
Молекулы разных ДНК отличаются друг от друга последовательностью расположения азотистых оснований в полинуклеотидной цепи. Никаких ограничений относительно последовательности нуклеотидов не существует, но, в силу правила Чаргаффа, последовательность оснований в одной цепи определяет их последовательность в другой.
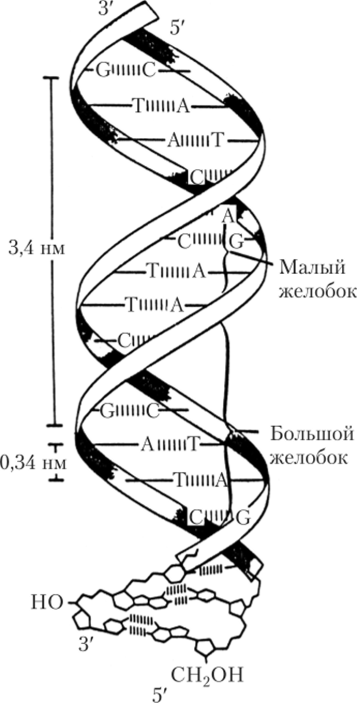
Рис. 25. Двойная спираль ДНК.
Цепи, образующие двойную спираль, удерживаются друг около друга за счет водородных связей между комплементарными основаниями. При этом аденин с тимином образуют две, а гуанин с цитозином — три водородные связи (см. рис. 2.5). Ширина спирали — 1,8 нм. Каждый виток спирали содержит 10 пн, расстояние между основаниями 0,34 нм, а длина полного витка спирали — 3,4 нм.
На поверхности спирали можно выделить большую и малую бороздки (желобки). Модель Уотсона — Крика описывает й-форму ДНК, наиболее распространенную в клетке. Тем не менее существуют и другие формы спирали ДНК.
Повышение температуры или изменение величины pH может привести к разрыву водородных связей и расплетанию двойной спирали ДНК, а при восстановлении прежних условий разошедшиеся цепи снова образуют двойную спираль. Процесс разделения цепей называется денатурацией, или плавлением, а процесс воссоединения — ренатурацией, или реассоциацией.
Из гипотезы двойной спирали следует, что информация, закодированная в ДНК, может быть точно воспроизведена, поскольку, согласно принципу комплементарности, каждая цепь ДЫК служит матрицей для образования новой комплементарной цепи. При этом правильность репликации каждой из цепей ДНК обеспечивается точным соотношением и стабильностью пар Г = ЦиА = Тв двух дочерних молекулах. Каждая из новых цепей содержит одну цепь родительской ДНК и новую комплементарную ей цепь.
Могут быть образованы гибриды ДНК, полученные из разных организмов, а также состоящие из РНК и одной из цепей ДНК. На этом свойстве нуклеиновых кислот основаны гибридизациопиые тесты, имеющие важное методическое значение. Они позволяют:
- 1) определять близость сравниваемых видов;
- 2) исследовать структуру генома;
- 3) изучать интенсивность экспрессии конкретного гена за счет ДНКРНК гибридизации;
- 4) идентифицировать индивидуальные гены.