Популяционный уровень экотоксических эффектов

Жизнеспособность семенного потомства в условиях химического загрязнения среды. Успешное прорастание семян при химическом загрязнении среды обусловлено рядом причин, связанных либо с условиями формирования полноценных семян, либо с качеством среды, влияющим на уже сформировавшиеся семена в процессе развития проростков. Среди последних важными являются тип загрязнения, продолжительность… Читать ещё >
Популяционный уровень экотоксических эффектов (реферат, курсовая, диплом, контрольная)
Небрежное отношение к биосфере ведет к нарушению ее правильной работы и означает, что люди не смогут вообще существовать на Земле.
Н. В. Тимофеев-Ресовски
Влияние токсических факторов на процессы воспроизводства в природных популяциях животных и растений.
Воспроизводство является одним из ключевых процессов, определяющих жизнеспособность природных популяций животных и растений. Особо значимы эти процессы в нестабильных условиях, в том числе при химическом загрязнении среды обитания, которое следует рассматривать в качестве одного из экстремальных факторов антропогенного происхождения.
Процессы воспроизводства в ценопопуляциях
Меристема, а также образующиеся из нее генеративные органы, в частности мужская генеративная сфера, являются одной из наиболее чувствительных к токсическому действию промышленных загрязнителей системой растений. При изучении реакции мужского гаметофита на воздействие внешних факторов принято различать оплодотворяющую способность (фертильность) и жизнеспособность пыльцы. Фертильность предполагает способность мужского гаметофита вызывать полное оплодотворение. Жизнеспособность пыльцы рассматривается как ее способность к росту на соответствующих тканях пестика.
На процессы микроспорогенеза, прорастания пыльцы и оплодотворения у растений влияет множество абиотических факторов: неблагоприятная погода, недостаток минерального питания, воздействие высоких или низких температур, высокие дозы химических веществ, радиоактивное загрязнение. Реакция пыльцы на действие физических и химических факторов может быть различна. Рядом исследователей установлено, что микроэлементы и УФ-радиация в малых дозах могут стимулировать ростовые процессы, а при высоких воздействиях — их подавляют.
Проведенные исследования по древесным и кустарниковым растениям свидетельствуют о негативном влиянии высоких концентраций тяжелых металлов и радиации на функциональное состояние и физиологическую активность пыльцы. Следствием подобного влияния могут стать замедленное прорастание пыльцы, нарушение роста пыльцевых трубок и полная остановка их роста, потеря жизнеспособности половых клеток (Третьякова и др., 2004; Beda, 1982).
Имеются сведения о влиянии высоких концентраций тяжелых металлов и радиационного излучения на функциональное состояние и физиологическую активность пыльцы травянистых растений (Бессонова, 1992; Попова и др., 1992; Реакция мужского гаметофита…, 2007; Третьякова и др., 2004; Holub et al., 1988).
Под действием химического загрязнения наблюдается уменьшение размеров пыльцевых зерен, что может рассматриваться в качестве одного из показателей адаптированности растений к техногенным условиям среды. Многие исследователи указывают на то, что под действием внешних факторов происходит образование деформированных (сморщенных, разрушенных, линзовидных, гигантских) и недоразвитых пыльцевых зерен (Бессонова, 1992; Сафонов, 1999; Чукпарова, 2001; Третьякова и др., 2004; Beda, 1982).
В условиях загрязнения среды промышленными поллютантами и тяжелыми металлами (Fe, Си, Mn, Cr, Zn и др.) отмечается резкое снижение оплодотворяющей способности пыльцы. В популяциях растений, находящихся в зонах антропогенного воздействия, повышается стерильность пыльцы, которая может достигать 50—80%, что обусловлено редукцией клеток археспория, цитопатологией генеративной ткани или нарушением мейоза (О возможностях…, 1996; Оценка последствий…, 1993).
Рассмотрим особенности функционального состояния мужского гаметофита (на примере форм одуванчика лекарственного Taraxacum officinale Wigg. s.l.).
Размеры пыльцевых зерен.
Отмечено уменьшение средних размеров пыльцы с увеличением химической нагрузки (рис. 7.1). Влияние химического загрязнения на исследуемый признак статистически значимо только в случае f. pectinatiforme (rs = -0,96; df = 7; р 0,001; для f. dahlstedtii rs = -0,18; df = 7; p = 0,7). На максимально загрязненном участке у растений f. dahlstedtii пыльца крупнее, чем у f. pectinatiforme. Различия между формами по исследуемому показателю подтверждены однофакторным дисперсионным анализом (F (1; 348) = 6,82; р
Уменьшение размеров пыльцевых зерен в условиях техногенного загрязнения показано также в работах В. П. Бессоновой (1992), А. У. Чукпаровой (2001) и др. Изменение величины пыльцы связано с нарушением процесса роста и деления клеток при формировании первичных клеток археспория и при формировании тетрады микроспор (Бессонова, 1992). Уменьшение размеров пыльцы некоторыми авторами рассматривается в качестве одного из показателей адаптированное™ растений к техногенным условиям среды (Лебедева, 1993).
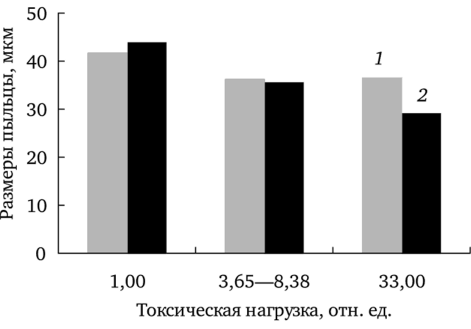
Рис. 7.1. Зависимость размеров пыльцы f. dahlstedtii (7) и f. pectinatiforme (2) от уровня химического загрязнения Анализ вариабельности этого показателя позволил установить, что влияние химического загрязнения проявляется в элиминации особей с крупными пыльцевыми зернами (рис. 7.2). Сравнение выборочных распределений растений с разными размерами пыльцы из ценопопуляции фонового участка с остальными показало статистически значимые различия.
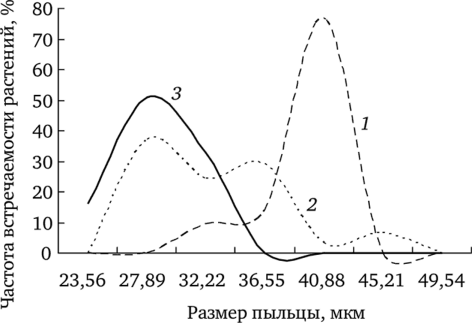
Рис. 7.2. Частота встречаемости растений с различными размерами пыльцы (по: Реакция мужского гаметофита…, 2007):
1 — фоновый; 2 — буферный; 3 — импактный участки С ростом химического загрязнения доля фертильной пыльцы снижается. Соответственно, на загрязненных участках при уменьшении фертильной пыльцы у исследуемого вида возрастает доля стерильных пыльцевых зерен.
Известно, что Т. officinale s.l. является видом с апомиктическим способом размножения. Вместе с тем высокие значения стерильности пыльцы у растений из ценопопуляций максимально загрязненных территорий по сравнению с фоновыми указывают на существенную роль химического загрязнения на процессы микроспорогенеза.
Оплодотворяющая способность пыльцы
Т. officinale s.l. в градиенте токсической нагрузки. Возникновение частичной или полной стерильности у растений часто связывают с нарушением правильности развития и строения пыльцы (Левина, 1981). В модели пошагового дискриминантного анализа показатель фертильности пыльцы обеих форм в нашем случае достоверно различается на всех участках (рис. 7.3).
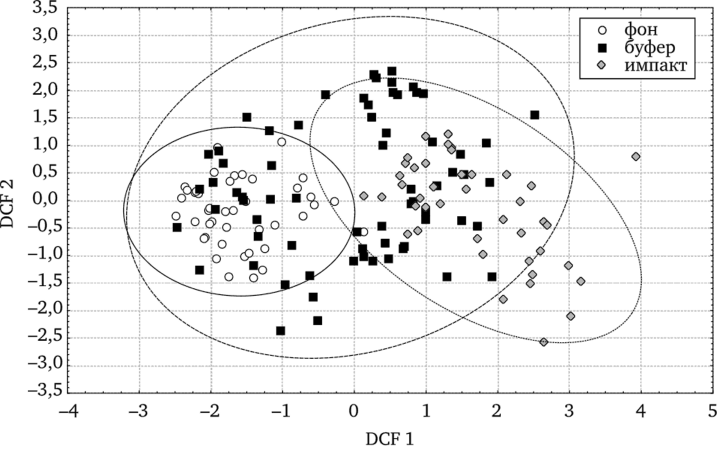
Рис. 7.3. Дискриминантный анализ фертильности пыльцы у ценопопуляций Т. officinale (по: Реакция мужского гаметофита…, 2007).
Установлены значительные различия по количеству фертильной пыльцы на одно растение вдоль первой дискриминантной канонической функции между ценопопуляциями минимально и максимально загрязненных участков. Буферные территории занимают промежуточное положение. Различия между всеми выборочными центроидами статистически достоверны (р < 0,05).
С ростом техногенного загрязнения доля фертильной пыльцы снижается (рис. 7.4). Соответственно, возрастает доля стерильных пыльцевых зерен.
В полной мере поражение биологических систем определяется не столько изменением средних значений показателей, сколько долей особей, имеющих максимально выраженный эффект воздействия. Условно можно принять в качестве такого критического значения (Скр) уровень фертильности пыльцы, который встречается менее чем у 5% особей фоновой выборки (рис. 7.5).
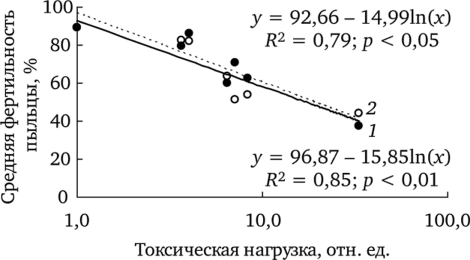
Рис. 7.4. Зависимость фертильности пыльцы f. dahlstedtii (7) и f. pectinatiforme (2) от уровня химического загрязнения (по: Реакция мужского гаметофита…, 2007).
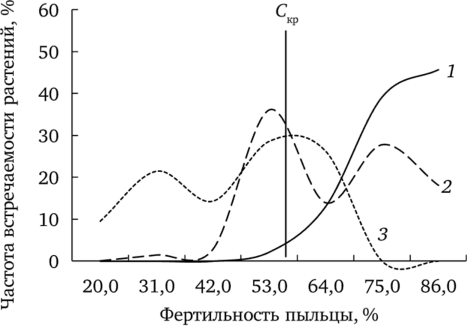
Рис. 7.5. Частота встречаемости растений f. dahlstedtii с различным уровнем фертильности пыльцы (по: Реакция мужского гаметофита…, 2007):
1 — фоновый; 2 — буферный; 3 — импактный участки Частота встречаемости растений обеих форм одуванчика с фертильностью пыльцы ниже обозначенного уровня изменяется от минимальной на фоновом участке до максимальной на импактном (рис. 7.6). Доля растений f. dahlstedtii и f. pectinatiforme с фертильностью пыльцы ниже критического уровня в выборках с фонового, буферного и импактного участков значимо различается (р < 0,001). Отметим, что в ценопопуляции импактного участка доля растений, имеющих эффект поражения, больше у f. pectinatiforme, чем у f. dahlstedtii (критерий Фишера, F (1; 138) = 18,55; р < 0,001).
Известно, что Т. officinale s.l. является видом с апомиктическим способом размножения. Вместе с тем высокие значения стерильности пыльцы из ценопопуляций максимально загрязненных территорий по сравнению с фоновыми указывают на существенную роль химического загрязнения в процессе формирования фертильной пыльцы.
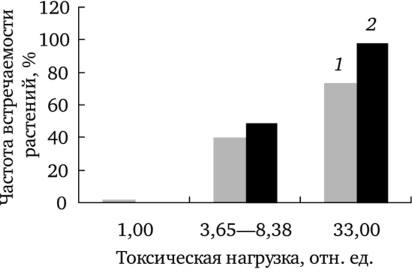
Рис. 7.6. Частота встречаемости растений f. dahlstedtii (7) и f. pectinatioforme (2) с фертильностью пыльцы ниже критического уровня.
Жизнеспособность пыльцы в условиях химического загрязнения среды. Наиболее точным методом определения жизнеспособности пыльцы является проращивание последней на рыльце пестика. Установлено, что пыльца одуванчика потенциально способна к оплодотворению, но часто при химическом загрязнении среды эта функция не реализуется. Поскольку Т. officinale s.l. является апомиктом, в условиях токсического загрязнения среды состояние мужского гаметофита, вероятно, не определяет благополучие ценопопуляций и может рассматриваться лишь в качестве диагностического признака.
Семенная продуктивность растений. Семенное возобновление вида — это поддержание оптимальной плотности популяции за счет семенного размножения. Успешность этого этапа определяет биологическую результативность всех предшествующих ему репродуктивных процессов.
Одним из факторов, определяющих популяционный уровень репродукции растений, является общая плотность популяции. При ее изменении у отдельных особей могут изменяться размер, высота, число генеративных побегов. При этом отдельные репродуктивные структуры особей реагируют на плотность по-разному: одни изменяются слабо, другие — высокопластичны. Репродуктивный процесс зависит также от возрастной структуры популяции.
Различают следующие показатели семенной продуктивности:
- • потенциальную продуктивность (ПСП) — количество семяпочек на один генеративный побег или одну особь;
- • реальную продуктивность (РСП) — количество зрелых полноценных семян на один генеративный побег или особь.
Изучению семенной продуктивности растений в условиях техногенного загрязнения среды посвящено значительное число работ (Васфилов и др., 1990; Влияние промышленного…, 1990; Использование…, 2002; Лянгузова и др., 1996; Савинов, 1998; Фролова, 1998; и др.). Характерно, что однозначного ответа репродуктивной структуры на химический пресс нет. У одних видов наблюдается снижение семенной продуктивности, массы и доли развитых семян, у других эти показатели остаются неизменными, а у третьих (многолетние травянистые растения) возможно даже повышение семенной продуктивности особей с одновременным повышением массы и качества семян. Приведем примеры реакции растений на химическое загрязнение среды.
В зоне действия медеплавильного комбината на Урале было зафиксировано уменьшение длины шишек сосны в среднем на 16—19%, их веса — на 37—50%, а веса семян — на 12—15%. В исследованиях Е. А. Мазной и И. В. Лянгузовой (1995) показано, что химическое загрязнение приводит к уменьшению количества крупных семян и увеличению мелких в одной ягоде черники, следствием чего является снижение массы 1000 шт. семян. Аналогичные результаты были получены Н. И. Ставровой (1990) для семян сосны обыкновенной.
Наряду с этим встречаются работы, в которых отмечено улучшение показателей репродуктивной сферы. У Taraxacum officinale показан рост семенной продуктивности в интенсивно загрязняемых биотопах при одновременном увеличении доли недоразвитых семянок в корзинке (Савинов, 1998). Показано также увеличение массы семян в условиях загрязнения (Аникеев, 1997). Повышенную семенную продуктивность в условиях загрязнения среды можно рассматривать в качестве проявления одного из приспособлений ценопопуляций к условиям токсического стресса.
Кроме влияния на семеношение, серьезное воздействие атмосферные загрязнения оказывают на прорастание семян и формирование подроста. В условиях загрязнения показано снижение энергии прорастания и всхожесть семян для сосны обыкновенной (Pinus sylvestris L.) (Веселкин, 2004; Черненькова, 2002), ели сибирской (Picea sibirica L.) (Черненькова, 1987), пырея ползучего (Elytrigia repens (L.) Nevski), звездчатки средней (Stellaria media L.), ежовника обыкновенного (Echinochloa crus-galli (L.) Beauv.) (Киреева и др., 2001), двукисточника тростниковидного (Phalaroides arundinacea (L.) Rausch.) (Семериков и др., 1990), яблони Палласа (MaZus pallasiana Jus.) (Плескач и др.,.
- 2002), одуванчика лекарственного (Taraxacum officinale Wigg.) (Пестова и др., 2004; Стволинская, 2000; Фролова, 1998), пшеницы (Triticum L.), овса (Avena L.), люцерны (Medicago L.) (Сысо и др., 2002), черники (Vaccinium myrtillus), брусники (Vaccinium vitis-idaea) (Лянгузова и др.,
- 2003), водяники гермафродитной (Empetrum hermaphroditum Hagerup) (Лянгузова и др., 1996), березы повислой (Betula pendula Roth.) (Васфилов и др., 1990). В вегетационных опытах по проращиванию семян Vaccinium myrtillus и Vaccinium vitis-idaea, сформировавшихся при различном уровне загрязнения среды отходами цинкоплавильного завода, всхожесть семян снижалась до 8—15 раз на сильно загрязненных тяжелыми металлами почвенных субстратах. Семенное потомство, собранное в зоне сильного техногенного загрязнения, помимо низкой энергии прорастания может выражаться задержкой темпов онтогенетического развития (Трубина, 2001).
Таким образом, в большинстве исследований отмечено ухудшение параметров генеративной сферы под действием промышленных загрязнителей, что, с одной стороны, связано с ухудшением общего жизненного состояния особей, с другой — с непосредственным поражением токсикантами репродуктивных органов.
С другой стороны, в условиях хронического загрязнения среды промышленными токсикантами могут формироваться устойчивые популяции растений, способные давать не только большое число потомков, но и сильные проростки, которые развиваются даже при высоких концентрациях тяжелых металлов в почве. Так, особи кровохлебки лекарственной (Sanguisorba officinalis L.), выращенные из семян растений импактной зоны, развиваются на загрязненной почве лучше, чем проростки из семян растений фоновой территории (Хантемирова, 1997). Аналогичные результаты были получены для семенного потомства астры альпийской (Aster alpinus L.), лабазника шестилепестного (Filipendula hexapetala Gilib.), дрока красильного (Genista tinctoria L.), костреца безостого (Bromopsis inermis Leyss. Holub.), мари белой (Chenopodium album L.) (Башмаков и др., 2003; Растения в экстремальных…, 1983).
На участках с нелетальными (физиологическими) концентрациями тяжелых металлов в почве всхожесть семян может быть достоверно выше контрольной, а при сублетальных концентрациях — оставаться на уровне контроля. Например, показатели жизнеспособности потомства черники (Vaccinium myrtillus L.), голубики (У. uliginosum L.), брусники (У. vitis-idaea L.) (Комалетдинова и др., 2002; Лянгузова и др., 1996), березы пушистой (Betula pubescens Ehrh.) остаются постоянными независимо от уровня химического загрязнения (Васфилов и др., 1990). Высокая устойчивость семенного потомства растений техногенных экотопов к тяжелым металлам свидетельствует о достижении этими популяциями определенного уровня генотипической дифференциации, отражающейся в наследовании признаков повышенной толерантности.
Жизнеспособность потомства может существенно меняться не только в градиенте химического загрязнения, но и в разные вегетационные сезоны (Влияние промышленного…, 1990; Репродуктивные возможности…, 2002). М. Гоздалик с соавт. (Gozdalik et al., 1998), наблюдая за разными аспектами семеношения деревьев (жизнеспособность пыльцы, эффективность опыления, размеры шишек и качество семян), произрастающих в условиях различных концентраций воздушных поллютантов и пыли, установили, что снижение качества семян в зоне промышленных эмиссий не носит постоянный характер и проявляется только в некоторые годы на отдельных деревьях. При этом наибольшее снижение жизнеспособности семян и пыльцы отмечено в годы с неблагоприятными атмосферными условиями.
Кроме того, семенную продуктивность определяет состояние материнских растений. В качестве примера проведем анализ морфологических показателей генеративной сферы средневозрастных особей одуванчика лекарственного, произрастающего в градиенте токсической нагрузки.
Показано, что длина цветоносного побега изменяется от 340,43 до 531,37 мм, его диаметр — от 3,50 до 5,19 мм, диаметр корзинки у основания — от 10,30 до 14,76 мм, высота корзинки — от 13,76 до 25,24 мм. Все исследуемые признаки характеризуются низким уровнем изменчивости (Cv = 2—3%, в единичных случаях достигает 6%). Аналогичные результаты были получены А. Б. Савиновым (1998) при изучении отдельных морфологических критериев генеративной сферы Т. officinale Wigg. из биотопов с разным уровнем загрязнения. В работе показано, что длина генеративного побега, диаметр его у основания и вершины, диаметр цветоложа у растений во всех ценопопуляциях имеют очень близкие средние значения. На низкую изменчивость параметров генеративной сферы (например, параметров цветка у любого вида растений) указывали также в своей работе А. Р. Ишбирдин и М. М. Ишмуратова (2004).
Количество генеративных побегов у отдельных средневозрастных особей Т. officinale Wigg. в градиенте химического загрязнения варьирует от 3 до 30 шт. Встречаются единичные экземпляры одуванчика с числом генеративных побегов до 47—50 шт. на одно растение. Трехфакторным дисперсионным анализом подтверждено высокозначимое влияние токсической нагрузки на число генеративных побегов у одуванчика во все исследуемые вегетационные сезоны (Северюхина и др.,.
2004).
Наличие более высокопродуктивных по числу генеративных побегов растений на загрязненных участках вряд ли можно объяснить прямым влиянием загрязнения почв. Скорее, отмеченный факт свидетельствует об исходной гетерогенности ценопопуляций Т. officinale s.l., при которой на всех участках присутствуют особи, обладающие высокой продуктивностью. При этом на загрязненных участках сохраняется их большая доля за счет повышенной элиминации иных вариантов. Аналогичные данные получены для других видов.
Общее количество семян в корзинке. Анализ общего числа семянок в корзинке у исследуемых морфологических форм Т. officinale показал, что оно варьировало от 100 до 280 шт. Встречались единичные корзинки с числом семян 67 и 323 шт. Однако однонаправленного изменения данного признака в градиенте увеличивающейся токсической нагрузки не выявлено.
Количество выполненных семян в корзинке. Общее число семян в корзинке еще не дает полной оценки семенного воспроизводства, так как не все семена в ней могут быть качественными (полноценными). Значение имеет число выполненных семян, так как именно эта фракция участвует в возобновлении популяции. Средние значения данного показателя варьируют от 55 до 210 шт. Химическое загрязнение приводит к снижению числа полноценных семянок на одном генеративном побеге по мере увеличения токсической нагрузки.
Выявлена зависимость между числом выполненных семян и их общим количеством в корзинке (рис. 7.7).
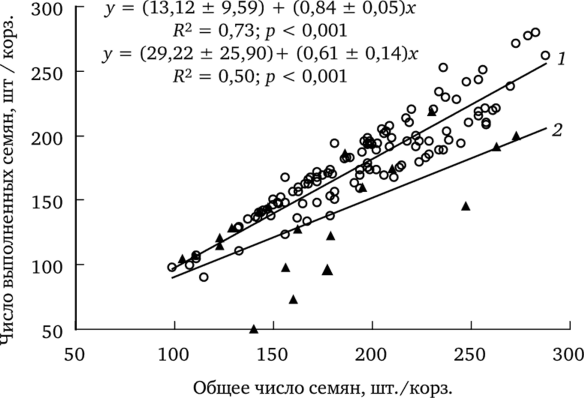
Рис. 7.7. Зависимость количества выполненных семян в корзинке от их общего числа (по: Жуйкова, 2009):
1 — фоновый (объединенный данные для двух форм) и импактный (f. dahlstedtii) участки; 2 — импактный участок (f. pectinatiforme).
В выборке с фонового участка различия между f. dahlstedtii и f. pectinatiforme по числу выполненных семян в корзинке статистически незначимы, что позволило объединить эти данные. У обеих форм одуванчика с этого участка увеличение числа семян в корзинке сопровождается ростом числа выполненных. Аналогичная зависимость показана для растений обеих форм из ценопопуляции импактного участка.
Количество неполноценных семян в корзинке. По мере снижения числа полноценных семянок, естественно, возрастает доля невыполненных. Количество пустых семянок в корзинках варьирует от 0 до 240 шт. Увеличение доли недоразвитых семян по мере роста загрязненности территории показано в работах В. А. Осколкова (1998а, б), Н. И. Ставровой (1990), А. Б. Савинова (1998).
На рис. 7.8 представлено соотношение выполненных и невыполненных семян в корзинке у растений в градиенте токсической нагрузки. У f. dahlstedtii на участках фоновой и буферной зон с увеличением загрязнения доля полноценных семян возрастает, однако значительно снижается на участке с максимальным загрязнением. У f. pectinatiforme процент выполненных семян на импактном участке значительно ниже, чем на всех остальных, что подтверждено S-методом множественных сравнений Шеффе (р < 0,01). На участках фоновой и буферной зон количество полноценных семян у исследуемой формы достигает 90% и более.
Подобная картина сохраняется независимо от условий вегетационного сезона. Увеличение относительного числа невыполненных семян по градиенту загрязнения может свидетельствовать о повреждающем действии токсикантов на женские генеративные структуры (Влияние промышленного…, 1990).
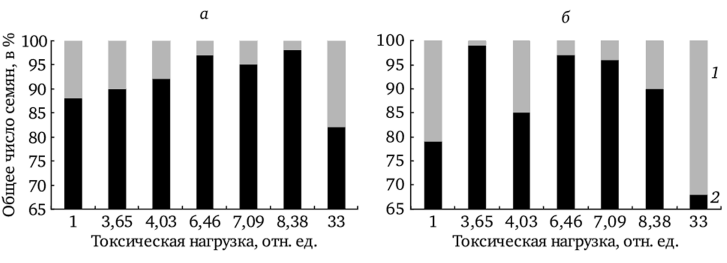
Рис. 7.8. Соотношение выполненных и невыполненных семянок в корзинке у f. dahlstedtii (а) и f. pectinatiforme (б) в градиенте токсической нагрузки:
1 — выполненных; 2 — невыполненных Средняя семенная продуктивность (ССП) растений напрямую зависит от количества генеративных побегов и от числа семян в корзинке. Выявлена положительная корреляция ССП с уровнем химического загрязнения почв тяжелыми металлами (рис. 7.9) (Репродуктивные возможности…, 2002; Северюхина и до., 2004). Аналогичное увеличение семенной продуктивности Т. officinale в условиях техногенного загрязнения показано в работе А. Б. Савинова (1998). В неблагоприятные вегетационные периоды ССП у растений обеих форм в условиях химического загрязнения остается на уровне фоновых.
Причину отмеченной закономерности объясняет сравнение частоты встречаемости растений с различной семенной продуктивностью на участках фоновой и импактной зон (рис. 7.10). Сравнение выборочных распределений растений с разной ССП из ценопопуляций фоновой и импактной зон показало статистически значимые различия между ними (f. dahlstedtii: у} = 48,29; df = 6; р < 0,001; f. pectinatiforme: X2 = 19,33; df — 6; p < 0,001). Можно предположить, что в ценопопуляции фоновой зоны представлен весь спектр растений, различающихся по этому показателю. Стрессовые воздействия, связанные с химическим загрязнением, приводят к элиминации растений с низкой продуктивностью, увеличивая долю высокопродуктивных.
Сложная внутрипопуляционная структура при воздействии внешних факторов стабилизирует популяцию как единую систему, обеспечивая необходимые в этих условиях репродуктивные возможности (Функциональное единство популяций, 1972).
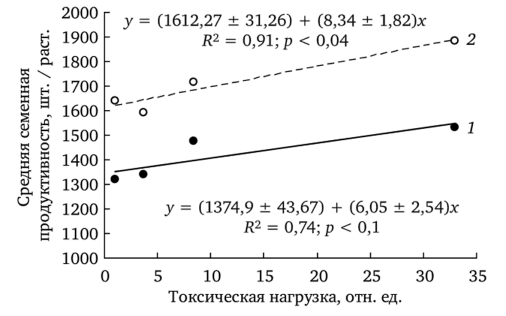
Рис. 7.9. Зависимость средней семенной продуктивности f. dahlstedtii (7) и f. pectinatiforme (2) от уровня токсической нагрузки (по: Безель и др., 2004).
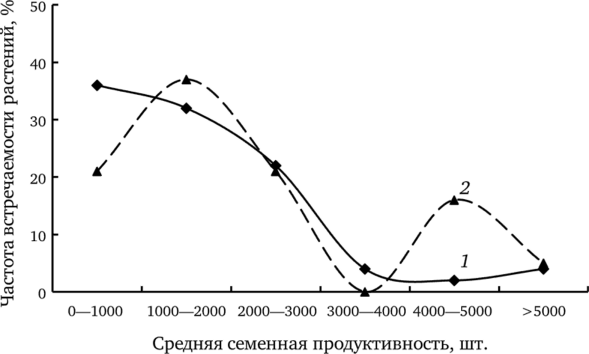
Рис. 7.10. Распределение средней семенной продуктивности f. pectinatiforme из ценопопуляций фоновой (7) и импактной (2) зон (по: Безель и др., 2004).
Реальная семенная продуктивность (РСП — число выполненных семян на растение) Т. officinale в среднем изменяется от 225 до 2430 шт. Анализ зависимости РСП от уровня токсической нагрузки показал, что в благоприятный год (гидротермический коэффициент (ГТК) сентября равен 2,3) наблюдается увеличение показателя у обеих форм одуванчика в градиенте химического загрязнения, что подтверждено методом регрессионного анализа (рис. 7.11).
В засушливый год (ГТК = 0,1) РСП у растений обеих форм с загрязненных участков остается на уровне фоновых.
Масса 1000 штук выполненных семян. Для более полной характеристики полноценной части урожая можно определить массу 1000 шт. выполненных семян. Установлено, что в выборках с этих участков наиболее часто встречаются растения с массой семян 0,28—0,35 г. По мере возрастания химического загрязнения на участках отмечено увеличение массы выполненных семян в корзинке (р < 0,01).
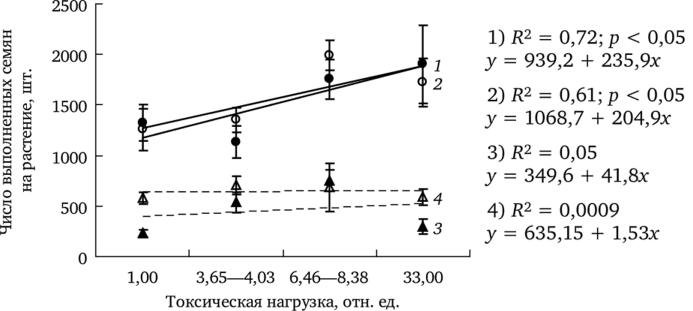
Рис. 7.11. Изменение реальной семенной продуктивности растений в градиенте токсической нагрузки (по: Жуйкова, 2009):
1,3 — f. dahlstedtii (ГТК = 2,3 и 0,1 соответственно); 2,4 — f. pectinatiforme (ГТК = 2,3 и 0,1 соответственно) Проведено сравнение частоты встречаемости особей с разной массой выполненных семян в условиях минимального и максимального уровня загрязнения (рис. 7.12). Наиболее часто встречаются растения с массой 0,32—0,40 г. Выборочные распределения не различаются (Х2 = 5,77; df = 6; р > 0,1). Однако ценопопуляция импактной зоны характеризуется модами в области более высоких значений массы выполненных семян. Доля растений с массой 1000 шт. семян более 0,57 г в этой ценопопуляции составляет 8%, а в фоновой выборке — 2% (критерий Фишера: F = 3,92; df = 1; 138; р < 0,05).

Рис. 7.12. Частота встречаемости растений с разной массой семян на участках фоновой (1) и импактной (2) зон (данные за 2000—2006 гг.).
(по: Жуйкова, 2009).
Жизнеспособность семенного потомства в условиях химического загрязнения среды. Успешное прорастание семян при химическом загрязнении среды обусловлено рядом причин, связанных либо с условиями формирования полноценных семян, либо с качеством среды, влияющим на уже сформировавшиеся семена в процессе развития проростков. Среди последних важными являются тип загрязнения, продолжительность воздействия, концентрация токсикантов, форма их соединения, а также индивидуальная устойчивость отдельных видов растений к высоким содержаниям токсикантов в почве. Рассмотрим, как изменяется жизнеспособность проростков Т. officinale, родительские растения которых произрастали в условиях разного химического загрязнения при культивировании их в дистиллированной воде и на загрязненных тяжелыми металлами почвах.
Всхожесть. Проведем сравнение результатов двух экспериментов (проращивание семян одуванчика лекарственного на дистиллированной воде и на почве) по всхожести — одному из показателей начальных этапов онтогенеза. В рулонной культуре в дистиллированной воде этот показатель в среднем равен 70%. На рис. 7.13 показано распределение доли проростков с различной выживаемостью в условиях минимального и максимального уровней химического загрязнения.
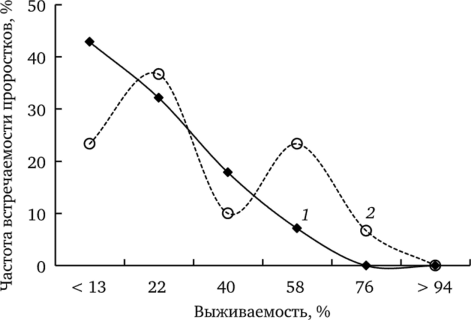
Рис. 7.13. Частота встречаемости проростков f. dahlstedtii с различной выживаемостью из ценопопуляций фоновой (7) и импактной (2) зон при проращивании на дистиллированной воде Большую долю в выборке фонового участка составляют проростки с низкой выживаемостью (10—58%). Среди семенного потомства импактной зоны наблюдается дифференциация по исследуемому показателю: наряду с проростками, имеющими низкую выживаемость, встречаются и проростки с высокой выживаемостью (> 58%). Сравнение выборочных распределений статистически значимо (%2 = 23,6; df= 5; р < 0,001).
В условиях почвенного фона всхожесть семян превышает 90% (рис. 7.14).
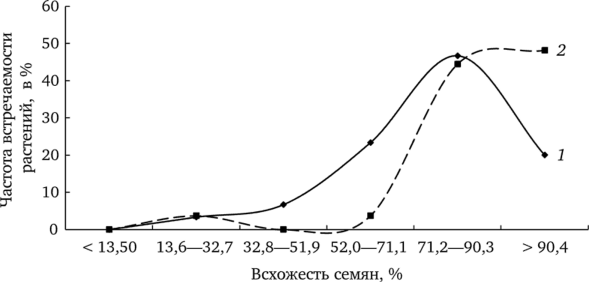
Рис. 7.14. Частота встречаемости растений с разной всхожестью семенного потомства при проращивании в условиях водного (7) и почвенного (2) фона.
Подобный характер вариационных кривых определяет более высокие значения всхожести семенного потомства одуванчика при проращивании в условиях почвенного фона. Аналогичная картина наблюдается и при анализе выживаемости одномесячных проростков.
Скорее всего, адаптация к повышенному содержанию тяжелых металлов в почве у растений загрязненной территории достигается путем дифференциации семенного потомства импактной ценопопуляции на особи с низкой и высокой выживаемостью. Аналогичные результаты были получены для семенного потомства астры альпийской, лабазника шестилепестного, дрока красильного, костреца безостого, потомства Chenopodium album L. (Башмаков и др., 2003).
Формирование корней проростков. Интересные данные получены при сравнении длины главного корня проростков Т. officinale при культивировании на загрязненной почве и в дистиллированной воде. Минимальные значения этого показателя у проростков во всех вариантах выявлены на фоновом участке. При этом длина корня при проращивании в дистиллированной воде меньше, чем у корней, культивируемых на почвах соответствующих участков (рис. 7.15). Вероятно, почва фоновой и буферной зон обладает стимулирующим эффектом вследствие оптимальных доз необходимых микроэлементов. Почва импактного участка, содержащая высокие концентрации металлов-загрязнителей, ингибирует ростовые процессы. Длина корней проростков в этом случае достоверно меньше, чем при проращивании тех же семян в дистиллированной воде (рис. 7.15).
Во многих исследованиях показано, что в условиях хронического загрязнения среды формируются устойчивые ценопопуляции, способные не только давать большое количество потомков, но и формировать сильные проростки, способные хорошо развиваться даже при высоких концентрациях тяжелых металлов в почве. Кровохлебка лекарственная, выращенная из семян растений импактной зоны, развивается на загрязненной почве лучше, чем проростки из семян растении чистой территории (Хантемирова, 1997). Аналогичные результаты представлены относительно потомства костреца безостого (Кортиков, 1996), а также в работе Н. В. Алексеевой-Поповой (1990) относительно семенного потомства астры альпийской, лабазника шестилепестного и дрока красильного.
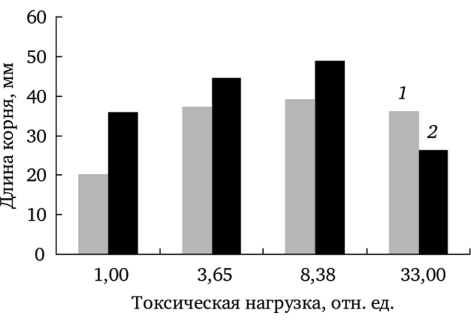
Рис. 7.15. Длина корней проростков Т. officinale, произрастающих на фоновых и загрязненных участках при проращивании в дистиллированной воде (7) и на почве с соответствующих территорий (2).
В табл. 7.1 приведены значения длины главного корня проростков T.officinale s.l. при проращивании в условиях почвенного фона.
Таблица 7.1
Средние значения ростовых критериев семенного потомства f. dahlstedtii (7) и f. rectinatipforme (2) в условиях почвенного фона (М ± т)
Токсическая нагрузка, отн. ед. | Длина корня, мм. | Число проростков с настоящим листом, % от выживших. | ||
1,00. | 33,06 ± 4,94. | 38,85 ± 4,81. | 87,01 ± 5,95. | 93,42 ± 6,41. |
3,65. | 39,14 ± 13,42. | 50,12 ± 4,15. | 74,26 ± 19,69. | 88,97 ± 5,17. |
8,38. | 45,12 ± 3,86. | 53,04 ± 10,84. | 97,27 ± 2,48. | 89,98 ± 19,83. |
33,00. | 27,12 ± 6,65. | 25,93 ± 5,88. | 82,08 ± 6,37. | 90,07 ± 4,97. |
Семенное потомство мари белой с импактной территории обладает повышенной всхожестью. На примере потомства голубки (р. Vaccinium) показано, что вследствие высокой жизнеспособности семян виды обладают достаточно высокими потенциальными возможностями к семенному размножению даже в условиях сильного промышленного загрязнения (Комалетдинова и др., 2002). Высокая устойчивость к тяжелым металлам семенного потомства растений химически деградированных экотопов свидетельствует о достижении этими популяциями определенного уровня генотипической дифференциации, проявляющейся в наследовании признаков повышенной толерантности.
Нарушения развития проростков. Высокие концентрации тяжелых металлов в окружающей среде не только ингибируют рост и развитие проростков, но и вызывают различные морфологические нарушения. В первую очередь действие токсикантов сказывается на корневой системе проростков. Следствием этого является резкое торможение роста и значительное уменьшение длины главного корня проростков. Корни растений в условиях сильного химического загрязнения укорочены и утолщены по всей длине, нередко наблюдаются их редукция, клубнеобразность, формирование скрученного укороченного гипокотиля. В результате нарушения функции корней, уменьшения их поверхности и постепенного отмирания резко снижается способность к поглощению питательных веществ.
По мере роста химического загрязнения участков увеличивается доля растений, имеющих некротические повреждения корневой системы, что является результатом токсического действия ионов тяжелых металлов и особенно четко проявляется при проращивании семенного потомства на почве (рис. 7.16).
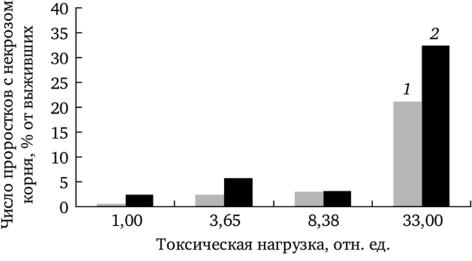
Рис. 7.16. Число проростков f. dahlstedtii (7) и f. rectinatipforme (2),.
имеющих некротические повреждения корешка при проращивании на почве (по: Северюхина, 2003).
Наряду с повреждением корневой системы наблюдается повреждение надземной части проростков: образование некротических и хлорофильных нарушений, деформация листьев и т. д.
Листообразование. Переход к автотрофному питанию в условиях почвенного фона идет в два раза интенсивнее по сравнению с водной рулонной культурой. Полученные различия объясняются присутствием в почве достаточного количества микроэлементов, необходимых для быстрого роста и развития проростков, и их отсутствием в дистиллированной воде.
Влияние условий вегетационного сезона на совокупность показателей жизнеспособности семенного потомства. В проблеме оценки влияния химического загрязнения на выживаемость семенного потомства большую роль играют погодные условия, при которых происходит формирование семенного потомства родительскими особями. Можно использовать метод дискриминантного анализа для оценки семенного воспроизводства и жизнеспособности семян, сформированных в условиях различного химического загрязнения в различные вегетационные сезоны. В качестве группирующей переменной в данном анализе использован год.
В первом случае в анализ были включены показатели семенной продуктивности (выборки 2000—2006 гг.): число цветоносов, общее число семян, число выполненных семян в корзинке, условно-реальная и реальная семенная продуктивность, масса 1000 выполненных семян. Во втором анализ проведен по совокупности показателей жизнеспособности потомства (выборки 2000—2004 гг.): энергии прорастания семян, всхожести, выживаемости проростков, листообразованию, длине корня. В качестве интегрального показателя использовали среднее значение расстояний Махаланобиса, рассчитанных для каждого года (рис. 7.17).
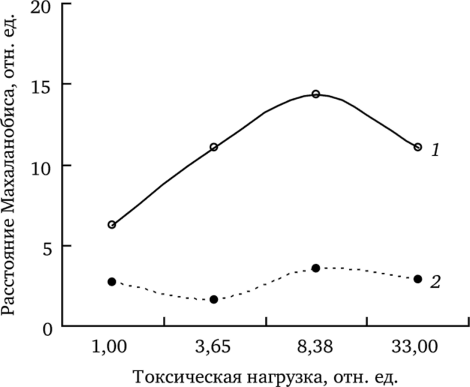
Рис. 7.17. Средние значения квадратов расстояний Махаланобиса, характеризующие близость показателей семенной продуктивности (7) и жизнеспособности потомства Т. officinale (2) в разные вегетационные сезоны (по: Жуйкова, 2009).
Минимальные различия между семенной продуктивностью разных лет для обеих форм показаны в выборке с фонового участка (рис. 7.17). В условиях среднего и высокого уровня загрязнения семенная продуктивность в разные годы различается значительнее. Следовательно, по показателям семенной продуктивности фоновая выборка менее чувствительна к условиям вегетационного сезона, чем буферная и импактная.
При анализе начальных этапов онтогенеза растений и ростовых критериев проростков в разные годы установлено, что различия между ними внутри фоновой, буферной и импактной выборок менее выражены (рис. 7.17). Средние квадраты расстояний Махаланобиса (MSD) — 2,7—2,9. Этот показатель лишь незначительно увеличивается на импактном участке.
Таким образом, семенная продуктивность растений в разные годы различается в большей степени, чем жизнеспособность потомства: средние квадраты расстояний Махаланобиса по показателям семенной продуктивности в выборках со всех участков выше, чем по показателям жизнеспособности. Это свидетельствует о меньшем влиянии изменяющихся условий вегетационных сезонов на жизнеспособность семенного потомства. Следовательно, показатели репродуктивной сферы материнских растений одуванчика в большей степени зависят от погодных условий в период формирования семенного потомства, чем показатели жизнеспособности уже сформированного ими потомства.
В заключение отметим, что воспроизводство растений определяется сочетанным действием химического загрязнения и погодно-климатических условий, в частности ГТК сентября предыдущего плодоношению года. От погодных условий в большей степени зависит семенная продуктивность растений, чем жизнеспособность потомства. Влияние условий вегетации материнских растений на жизнеспособность их потомства опосредованное. Возможно, основной отбор происходит на стадии семян, в результате которого во всех ценопопуляциях независимо от загрязнения и погодно-климатических факторов сохраняется определенная доля качественного потомства, способного обеспечивать устойчивость ценопопуляций. Совместное действие токсического и климатического факторов усиливает действие отбора.