Одномембранные органеллы.
Ботаника

Хлоропласт (рис. 85) — двумембранная пластида, в которой протекает фотосинтез за счет наличия пигментов: хлорофиллов, каротиноидов и ксантофиллов. Встречаются во всех зеленых органах растений, а также в зародышах части растений. Форма хлоропластов чаще всего линзовидная или эллипсоидальная. Являются самыми крупными пластидами, достигая в длину 4−10 мкм и 2−4 мкм в ширину, хорошо различимы… Читать ещё >
Одномембранные органеллы. Ботаника (реферат, курсовая, диплом, контрольная)
Аппарат Гольджи (комплекс Гольджи) состоит из отдельных диктиосом и пузырьков Гольджи (рис. 83). Диктиосомы представляют собой стопку плоских, не соприкасающихся друг с другом дисковидных цистерн, ограниченных мембранами. Часто диктиосомы переходят по краям в систему тонких ветвящихся трубок. Число диктиосом в растительной клетке обычно колеблется от одной до нескольких десятков. Пузырьки Гольджи отчленяются от краев диктиосомных пластинок или концов трубок и направляются обычно в сторону плазмалеммы или вакуоли. Считается, что в диктиосомах синтезируются полисахариды. Пузырьки Гольджи транспортируют образовавшиеся полисахариды, которые у растений нередко участвуют в формировании срединной пластинки и клеточной стенки. Кроме того, комплекс Гольджи, по-видимому, участвует в формировании вакуолей.
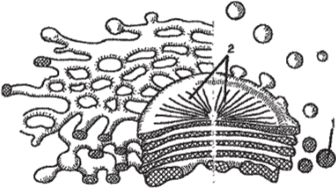
Рис. 83. Строение диктиосомы (аппарат Гольджи):
1 — пузырек Гольджи; 2 — цистерна диктиосомы в плане.
Лизосомы — органоиды, главным образом, грибов и животных, в последние годы найденные и в клетках растений. Они представляют собой мелкие (0,2−18 мкм) цитоплазматические пузырьки, окруженные мембраной. Образуются в комплексе Гольджи или в эндоплазматической сети. Содержат набор гидролитических ферментов. С их помощью осуществляется внутриклеточное пищеварение. Полагают, что функция лизосом — пополнение питательных веществ при временном их недостатке за счет использования резервов протопласта. Лизосомы могут также лизировать стареющие органоиды.
Сферосамы — мелкие тельца, размером от 0,2 до 1,3 мкм, первоначально окруженные биологической мембраной и содержащие специфические ферменты. Функция сферосом состоит в накоплении жира. Зрелая сферосома обычно представляет собой каплю жира, окруженную биологической мембраной или белковой оболочкой.
Мелкие сферические или эллипсоидальные органоиды размером от 0,2 до 1,5 мкм, окруженные одной мембраной, получили общее название микротелец. Наиболее известные из них — ппиоксисомы и пероксисомы.
Гпиоксисомы содержат ферменты, необходимые для превращения жиров в углеводы, что происходит во время прорастания семян. В них осуществляется цикл глиоксиловой кислоты.
Пероксисомы обнаружены с помощью электронного микроскопа в большинстве типов клеток. Функции пероксисом зависят от типа клетки. Подобно пластидам и митохондриям, пероксисомы до известной степени автономны. В ряде случаев они имеют непосредственное отношение к фотодыханию, играя важную роль в метаболизме гликолевой кислоты.
Двумембранные органеллы.
Митохондрии неотъемлемая часть почти всех живых эукариотических клеток (рис. 84). Округлые или цилиндрические, реже нитевидные органеллы, видимые в световой микроскоп. Длина их достигает 10 мкм, диаметр 0,2−1 мкм. Внутренняя мембрана образует выросты — кристы. Образование крист увеличивает внутреннюю активную поверхность. При аэробном дыхании на кристах происходят окислительное фосфорилирование и перенос электронов. Внутреннее содержимое митохондрий — матрикс — включает рибосомы (прокариотического типа), специфические иРНК и тРНК, кольцевую молекулу ДНК и фосфатные гранулы. В матриксе находятся ферменты, участвующие в цикле Кребса и окислении жирных кислот. Основная функция митохондрий — образование энергии.
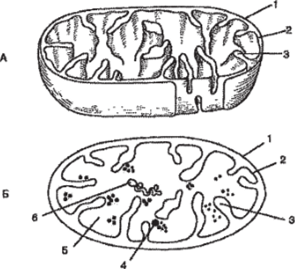
Рис. 84. Схемы строения митохондрии в трехмерном изображении (А) и на срезе (Б):
- 1 — наружная мембрана митохондрии; 2 — внутренняя мембрана; 3 — криста;
- 4 — рибосома; 5 — матрикс; 6 — кольцевая нить ДНК
Типы пластид. Эго органоиды, характерные исключительно для растительных клеток. Предшественниками пластид являются так называемые пропласгиды, мелкие обычно бесцветные образования, находящиеся в делящихся клетках корней и побегов. Форма, размеры, строение и функции пластид различны. По окраске (наличию или отсутствую пигментов) различают три типа пластид: зеленые хлоропласты, желто-оранжевые и красные хромопласты, бесцветные лейкопласты. Возможно взаимное превращение пластид. Обычно в клетке встречается только один тип пластид.
Хлоропласт (рис. 85) — двумембранная пластида, в которой протекает фотосинтез за счет наличия пигментов: хлорофиллов, каротиноидов и ксантофиллов. Встречаются во всех зеленых органах растений, а также в зародышах части растений. Форма хлоропластов чаще всего линзовидная или эллипсоидальная. Являются самыми крупными пластидами, достигая в длину 4−10 мкм и 2−4 мкм в ширину, хорошо различимы в световой микроскоп. Внутренняя среда хлоропласта — студенистообразный матрикс — строма. Строма содержит рибосомы, кольцевую молекулу ДНК и капельки масла. В строме протекает темновая фаза фотосинтеза, в которой непосредственно происходит синтез органических соединений с использованием энергии, синтезированной в световую фазу. Внутренняя мембрана вдается в полость хлоропласта выростами. Как строма, так и выросты внутренней мембраны формируют в полости хлоропласта сложную систему мембранных поверхностей, отграничивающих особые плоские мешки, называемые тилакоидами, или ламеллами. Группы дисковидных тилакоидов связаны друг с другом таким образом, что их полости оказываются непрерывными. Эти тилакоиды образуют стопки (наподобие стопки монет), или граны. Тилакоиды стромы (фреты) объединяют граны между собой. В мембранах тилакоидов сосредоточен главнейший пигмент зеленых растений — хлорофилл и вспомогательные пигменты — каротиноиды. В гранах осуществляется световая фаза фотосинтеза, в ходе которой осуществляются процессы циклического и нециклического фосфорилирования и фотолиза воды под действием квантов света. Хлоропласты могут превратиться в хромопласты (пожелтение листьев) или в лейкопласты (если поместить растение в темноту).
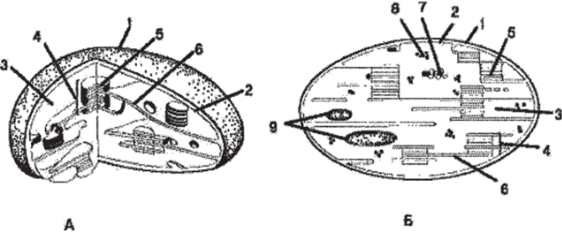
Рис. 85. Строение хлоропласта: А — объемная схема строения хлоропласта; Б — схема среза через хлоропласт: 1 — наружная мембрана;
- 2 — внутренняя мембрана; 3 — строма; 4 — грана; 5 — тилакоид внутренней мембраны (граны); 6 — тилакоиды стромы (фреты); 7 — ДНК;
- 8 — рибосомы хлоропласта (отличаются от цитоплазматических рибосом);
- 9 — крахмальные зерна
Хромопласт — окрашенная пластида, содержащая пигменты: каротиноиды (оранжевые) и ксантофиллы (желтые). Хромопласты являются конечным этапом в развитии пластид, поэтому у них, как правило, отсутствует внутренняя мембранная система. От хлоропластов они отличаются меньшими размерами и разнообразной формой (рис. 86). Больше всего хромопластов в плодах томата, красного перца, в цветках, где их яркая окраска служит для привлечения насекомых и птиц, участвующих в опылении растений и распространении семян.
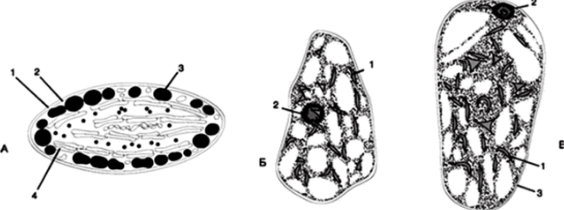
Рис. 86. Строение хромопласта: А: внешний вид: 1 — наружная мембрана;
- 2 — внутренняя мембрана; 3 — жировые капли; 4 — ламеллы; Б, В: хромопласты в клетках мякоти зрелых плодов рябины (Sorbus aucuparia) и боярышника (Crataegus sanguined) соответственно:
- 1 — хромопласты; 2 — ядро; 3 — стенка клетки
Лейкопласт — бесцветная пластида, не содержащая пигментов. В отличие от хлоропластов у лейкопластов слабо развитая мембранная система и редко расположенные одиночные тилакоиды. Лейкопласты могут превращаться в хлоропласты и хромопласты. Лейкопласты приспособлены для хранения запасных питательных веществ, поэтому их особенно много в запасающих органах: корнях, семенах, молодых листьях (рис. 87). В амилопластах откладывается запасной крахмал; в олеопластах (элайопластах) — липиды; в протеинопластах — белки.
Ядро, его строение, функции и значение в жизни клеток. Ядро — обязательная и существеннейшая часть живой клетки всех эукариотических организмов. Это место хранения и воспроизведения наследственной информации, определяющей признаки данной клетки и в конечном итоге всего организма в целом. Ядро также служит центром управления обменом веществ и почти всех процессов, происходящих в клетке. Из органоидов лишь митохондрии и пластиды в некоторой степени автономны и в части своих функций независимы от ядра. Клетки с удаленным ядром, как правило быстро погибают. В живых клетках ядро отсутствует в норме лишь в зрелых члениках ситовидных трубок флоэмы.
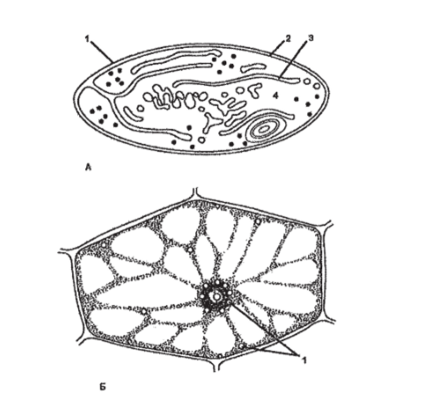
Рис. 87. Строение лейкопласта: А: внешний вид лейкопласта:
1 — наружная мембрана; 2 — внутренняя мембрана; 3 — ламелла; 4 — строма; Б: лейкопласты (1) в клетках листа традесканции Чаще всего в клетках эукариот имеется лишь одно ядро, редко — два или несколько (протоктисты). Нормальное длительное существование в одной клетке двух неслившихся ядер (дикарион) характерно для многих грибов.
Размеры ядра различны от 2−3 до 500 мкм (у половых клеток). Однако без специальной окраски ядро малозаметно. Форма его чаще всего шаровидная или эллипсоидальная. В молодых, особенно меристематических, клетках оно занимает центральное положение, но позднее обычно смещается к оболочке, оттесняемое растущей вакуолью.
Общий план строения ядра сходен у большинства эукариотических организмов. Снаружи оно окружено двойной мембраной — ядерной оболочкой, пронизанной порами, на краях которых наружная мембрана переходит во внутреннюю. Наружная мембрана ядерной оболочки в некоторых местах объединяется с эндоплазматической сетью. По-видимому, ядерная оболочка — специализированная часть этой сети.
Содержимое интерфазного неделящегося ядра составляет кариоплазма (или ядерный сок), близкая по структуре к гиалоплазме. В кариоплазму погружены оформленные элементы: хроматин (плотное вещество ядра, хорошо окрашиваемое основными красителями) и ядрышки, а также рибосомы. В процессе клеточного деления хроматин все более уплотняется и, в конце концов, собирается в хромосомы.
Число хромосом в ядре, так же как их размер и форма (хромосомный набор), постоянно для каждого вида организмов. В ядрах соматических (неполовых) клеток содержится диплоидный (двойной) набор хромосом — 2п. Он образуется в результате слияния двух половых клеток с гаплоидным (одиночным) набором хромосом — п. У твердой пшеницы в половых клетках по 14 хромосом, а в соматических — по 28. Диплоидный набор хромосом мягкой пшеницы — 42, значит, в ядре содержится 21 пара гомологичных хромосом. Совокупность признаков хромосомного набора, характерных для вида, получила название кариотип. Постоянство кариотипа каждого вида поддерживается в процессе деления. Иногда возникают ядра с набором хромосом, равным Зп, 4п и т. д. Их называют полиплоидными.
По химическому составу ядро отличается высоким содержанием ДНК. Большая часть ДНК клетки находится в ядре, в комплексах с ядерными белками. Основная масса ДНК сосредоточена в хроматине — особых нуклеопротеидных нитях, рассеянных по всему ядру.
В ядре заметно одно или несколько ядрышек. Подобно хроматину, ядрышки не имеют мембраны и свободно лежат в кариоплазме, состоя в основном из белка. Они содержат около 5% РНК и имеют большую плотность, чем ядро. Основная функция ядрышек — синтез некоторых форм РНК (в основном рибосомной) и формирование предшественников рибосом (субъединиц).
Вакуоли и клеточный сок. Клеточный сок образуется в процессе жизнедеятельности протопласта. Полости, заполненные клеточным соком и отграниченные тонопластом, называются вакуолями. Для большинства зрелых клеток характерна центральная крупная вакуоль, которая занимает 70−90% объема клетки. Она возникает при слиянии мелких цитоплазматических вакуолей, которые образуются цистернами эндоплазматической сети. В образовании вакуолей участвует и аппарат Гольджи, где изолируются продукты вторичного обмена, транспортируемые потом пузырьками Гольджи в вакуоль. Мембрана пузырька идет на пополнение тонопласта, а содержимое включается в состав клеточного сока. Один из путей образования вакуолей — изоляция участков цитоплазмы и последующее переваривание её в лизосомах.
Клеточный сок — слабо концентрированный водный раствор минеральных и органических соединений, образующих истинные и коллоидные растворы. При обезвоживании вакуолей они переходят в форму кристаллов или кристаллоидов. Клеточный сок имеет в основном слабокислую реакцию (pH 3−5). Химический состав его зависит от вида растения, его возраста и состояния.
Физиологическая роль веществ клеточного сока различна. В нём накапливаются и запасные питательные вещества (простые белки, углеводы), и вещества, регулирующие взаимовлияние растений, растений и животньгх (гликозиды, пигменты, алкалоиды), и осмотически деятельные соединения (соли органических и неорганических кислот).
Функции вакуолей заключаются, с одной стороны, в накоплении запасных и «захоронении» отбросов, то есть конечных продуктов метаболизма клетки. Иногда вакуоли разрушают токсичные или ненужные клетке вещества. Обычно это выполняется специальными небольшими вакуолями, содержащими соответствующие ферменты. Такие вакуоли получили название лизосомных. Другая функция вакуолей — поддержание тургора и регуляция водно-солевого обмена. Между клеточным соком, протопластом и клеточными стенками постоянно происходит движение веществ и воды. Тонопласт легкопроницаем для воды и, обладая избирательной проницаемостью, замедляет выход из вакуоли ионов и сахаров. Поэтому при достаточном обводнении клеточных стенок вода будет поступать в вакуоль за счет диффузии. Однонаправленный процесс диффузии воды через полупроницаемую мембрану называется осмосом. Если клеточный сок имеет более высокую концентрацию, то вода будет проникать в вакуоль. Увеличиваясь при этом в объеме, вакуоль будет давить на цитоплазму, прижимая её к клеточной стенке и создавая тургорное давление. Клеточная стенка в силу своей упругости будет оказывать обратное давление на протопласт. Это противодавление клеточных стенок называется тургорным натяжением. По мере поступления воды в клетку оно возрастает.
Поступление воды в клетку, хотя и происходит на основе осмоса, лимитировано присутствием ограниченно растяжимой клеточной стенки. Когда будет достигнут предел растяжимости клеточной стенки, всасывание воды прекратится. Концентрация клеточного сока будет наименьшей, тургорное натяжение — максимальным, клетка имеет наибольший возможный объем. Напряженное состояние клеточной стенки, создаваемое гидростатическим давлением внутриклеточной жидкости, называется тургором.
Тургор — нормальное физиологическое состояние растительной клетки. Благодаря тургору растение сохраняет свою форму, занимает определенное положение в пространстве, противостоит механическим воздействиям. Если в клетку в состоянии тургора поместить в раствор, осмотическое давление которого выше, чем клеточного сока, то вода устремится в сторону более концентрированного раствора и будет выходить из клетки. Сокращение объема вакуоли приведет к уменьшению давления её на цитоплазму, а цитоплазмы — на клеточные стенки. Клеточные стенки в силу своей эластичности станут менее растянутыми, а объем клетки уменьшится. Если объем клетки достигнет минимума, а уменьшение объема цитоплазмы будет продолжаться, то, сжимаясь, она начнет отставать от стенок и постепенно соберется в центре клетки. Наступает плазмолиз — состояние, обратное тургору. Если плазмолизированную клетку поместить в чистую воду, то тургор восстановится, произойдет деплазмолиз. Длительный и сильный плазмолиз может вызвать гибель клетки, при частичном плазмолизе растение увядает.
Отличие растительной клетки от грибной и животной клеток. Характерные признаки и сравнительное рассмотрение представлено в таблице 2.
Таблица 2.
Отличие растительной клетки от грибной и животной клеток.
Признак. | Растительная клетка. | Грибная клетка. | Животная клетка. |
Наличие оформленного ядра. | |||
Количество ядер | От 1. до нескольких. | ||
Наличие клеточной оболочки. | ; | ||
Основной компонент клеточной стенки. | Целлюлоза. | Хитин. | ; |
Наличие пластид. | ; | ; | |
Наличие центриолсй. | ; | Есть не у всех. | |
Наличие вакуолей. | Крупная центральная вакуоль. | Центральная вакуоль. | Нет крупных вакуолей (суммарный объем не превышает 5% от всего объема клетки). |
Запасное вещество. | Крахмал. | Гликоген. | Гликоген. |
Тип питания. | Автотрофный. | Гетеротрофный. | Гетеротрофный. |
Подвижность. | Неподвижны. | Неподвижны. | Подвижны. |
Клеточная оболочка и ее видоизменения. Образование вторичной оболочки, типы пор. Клеточная оболочка (клеточная стенка) — это структурное образование, располагающееся по периферии клетки, за пределами плазмалеммы, придающее клетке прочность, сохраняющее её форму и механически защищающее протопласт.
Клеточная стенка растений противостоит высокому осмотическому давлению большой центральной вакуоли и препятствует разрыву клетки. Кроме того, совокупность прочных клеточных стенок выполняет роль своеобразного внешнего скелета, поддерживающего форму растения и придающего ему механическую прочность. Клеточная стенка, обладая большой прочностью, в то же время способна к росту, и прежде всего к росту растяжением. Эти два в известной степени противоположных требования удовлетворяются за счет особенностей её строения и химического состава. Клеточная стенка, как правило, прозрачна и хорошо пропускает солнечный свет. Через неё легко проникает вода и низкомолекулярные вещества, но для высокомолекулярных веществ она полностью или частично непроницаема.
Стенка клетки представляет собой продукт жизнедеятельности её протопласта. Поэтому стенка может расти, только находясь в контакте с ним. Однако при отмирании протопласта стенка сохраняется, и мертвая клетка может продолжать выполнять функции проведения воды или играть роль механической опоры.
Основу клеточной стенки составляют высокополимерные углеводы, молекулы целлюлозы (клетчатки), собранные в сложные пучки — фибриллы, образующие каркас. Молекулы целлюлозы состоят из большого числа линейно расположенных мономеров — остатков глюкозы. Целлюлоза очень стойка, не растворяется в разбавленных кислотах и даже в концентрированных щелочах. Из 30 млрд т углерода, которые высшие растения ежегодно превращают в органические соединения, около Уг приходится на целлюлозу. Целлюлозный скелет придаёт клеточной стенке механическую прочность. Первоначально число микрофибрилл, образованных молекулами целлюлозы, в клеточной стенке относительно невелико, но с возрастом оно увеличивается, и клетка теряет способность к растяжению. Реактивом на целлюлозу является хлор-цинк-йод (Cl-Zn-I), дающий сине-фиолетовое окрашивание. Целлюлозный каркас клеточной стенки заполнен нецеллюлозными молекулами матрикса. В состав матрикса входят полисахариды, называемые гемицеллюлозами; пектиновые вещества (пектин), очень близкие к гемицеллюлозам, и гликопротеиды. Пектиновые вещества, сливаясь между соседними клетками, образуют срединную пластинку, которая располагается между первичными оболочками соседних клеток. При растворении или разрушении срединной пластинки (что происходит в мякоти созревших плодов) возникает мацерация (от лат. maceratio — размягчение). Естественную мацерацию можно наблюдать у многих перезрелых плодов (арбуз, дыня, персик). Искусственную мацерацию (при обработке тканей щелочью или кислотой) используют для приготовления различных анатомических и гистологических препаратов.
Клеточная стенка слоиста и состоит из одной или нескольких оболочек. При делении меристематических клеток первоначально формируется срединная пластинка, образованная главным образом аморфными пектиновыми веществами. Протопласт каждой дочерней клетки откладывает со своей стороны на срединную пластинку собственную первичную оболочку, состоящую главным образом из пектиновых веществ и гемицеллюлоз. При этом клетка растет, растягиваясь в основном под влиянием тургорного давления, а новые порции гемицеллюлоз «внедряются» в растягивающуюся оболочку. Оболочки делящихся и растущих клеток называют первичными. Они богаты водой, и содержание целлюлозы в них относительно невелико (не более 30%). Тонкие участки первичной оболочки называются первичными поровыми полями.
Для многих клеток отложение новых слоев оболочки прекращается с прекращением роста клетки. У других клеток отложение оболочки изнутри продолжается и по достижении окончательного размера. При этом толщина клеточной стенки увеличивается за счет наложения, а объем полости клетки сокращается. Такой процесс носит название вторичного утолщения клеточной стенки, а сама оболочка называется вторичной. Вторичная оболочка выполняет главным образом механическую функцию. Наиболее обычна она в клетках опорных тканей. Химический состав вторичной оболочки иной, чем у первичной. В ней содержится меньше воды, а количество целлюлозы достигает 40−50% от массы сухого вещества. Вторичная оболочка иногда откладывается неравномерно. У части сосудов она имеет вид отдельных колец или спиралей. Это позволяет клеткам сохранить способность к растяжению в длину. Стенки клеток, имеющие вторичные утолщения, часто одревесневают вследствие отложения в их матриксе лигнина.
Клеточная стенка в процессе жизнедеятельности может подвергаться различным видоизменениям — одревеснению, опробковению, ослизнению, кутинизации, минерализации (табл. 3).
Одревеснение клеточной стенки связано с внедрением между молекулами целлюлозы лигнина, который является самым распространенным (после целлюлозы) полимером растительных клеток. Он увеличивает жесткость оболочки, вызывая одревеснение клеточных стенок; лигнин обычно содержится в клетках, выполняющих опорную или механическую функцию. Реактив на лигнин НС1 (концентрированная) + флороппоцин дает красно-малиновое окрашивание.
Таблица 3.
Характерные изменения вторичной клеточной стенки.
Изменение. | Вещества, вызывающие изменения. | Реактив. | Результат. |
Наслаивание целлюлозы. | Целлюлоза. | Cl-Zn-I. | Сине-фиолетовое окрашивание. |
Кутинизация. | Кутин. | Судан III. | Оранжевое окрашивание. |
Одревеснение. | Лигнин. | Флороглюцин+кислота хлористоводородная концентрированная. | Красно-малиновое окрашивание. |
Опробковение. | Суберин. | Судан III. | Оранжевое окрашивание. |
Минерализация. | Кремнезем, соли Са, Mg и др. | Сжигание. | Минеральный остаток. |
Ослизнение. | Углеводы — слизи, камеди. | Вода. | Ослизнение. |
Кутин, суберин и воск — жироподобные вещества. Кутин и воск обычно откладываются на поверхности клеток эпидермы. Кутиновая пленка образует кутикулу Суберин пропитывает клеточные стенки вторичной покровной ткани, вызывая опробковение. В момент завершения опробковения протопласт клетки разрушается, а клеточная стенка пробки становится непроницаемой для воды и газов. Кутин и суберин встречаются в комбинации с восками и предотвращают чрезмерную потерю воды растением и проникновение в его клетки различных бактерий и грибов.
Клеточные стенки могут пропитываться оксалатами и кремнеземом, что придает им твердость и хрупкость и приводит к их минерализации. Отложение кремнезема характерно для стеблей и боковых побегов хвощей, стеблей злаков и осок.
Некоторые клеточные стенки кожуры семян (льна) способны к ослизнению. Это происходит за счет превращения целлюлозы и пектина в слизи и камеди, которые, являясь полимерами, сильно набухают при соприкосновении с водой. Слизи удерживают влагу, защищая семена от высыхания, и закрепляют их на определенном месте, с частицами почвы. Например, у стеблей вишни выделение слизей и камедей наблюдается из пораненных участков. Камедь при этом выделяется в виде наплывов (вишневый клей); этот процесс называют гуммозом.
В стенках соседних клеток, как правило, одна против другой, образуются поры. Они чаще всего закладываются там, где есть первичные поровые поля. Порами называют отверстия во вторичной оболочке, где соседние клетки разделяют лишь первичная оболочка и срединная пластинка. Участки первичной оболочки и срединную пластинку, разделяющие соседствующие поры смежных клеток, называют поровой мембраной, или замыкающей плёнкой поры. Замыкающую плёнку поры пронизывают плазмодесменные канальцы, но сквозного отверстия в порах обычно не образуется (рис. 88).
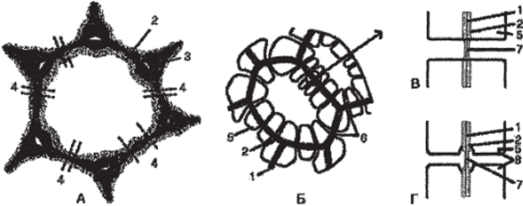
Рис. 88. Первичные поровые поля, поры и плазмодесмы:
А — паренхимная клетка с первичной оболочкой и первичными поровыми полями; Б — клетки с вторичными оболочками и многочисленными порами;
В — пара простых пор; Г — пара окаймленных пор; 1 — срединная пластинка;
- 2 — первичная оболочка; 3 — межклеточное пространство; 4 — плазмодесмы в первичном поровом поле; 5 — вторичная оболочка; 6 — поры;
- 7 — поровая мембрана; 8 — окаймление
Содержимое соседних клеток связано друг с другом через специальные цитоплазматические тяжи — плазмодесмы. Плазмодесмы располагаются в плазмодесменных канальцах поровой мембраны. Посредством плазмодесм осуществляется пересдача раздражений и активное передвижение некоторых веществ от клетки к клетке. Каждая пора имеет поровую камеру. В тех случаях, когда откладывается мощная вторичная оболочка, камеры превращается в узкие поровые каналы. В клетках паренхимных и механических тканей вторичная оболочка обычно резко прерывается у краев камеры или порового канала, диаметр которых благодаря этому почти не изменяется по всей толще вторичной оболочки. Поры такого типа называются простыми, а комбинация двух простых пор — простой парой пор. В водопроводящих элементах — сосудах и трахеидах — вторичная оболочка нередко нависает над камерой в виде свода, зрительно образуя окаймление (рис. 89). Такие поры получили название окаймленных, или окаймленной парой пор. Поровая камера, ограниченная окаймлением, открывается в полость клетки через отверстие в окаймлении — апертуру поры. Поры облегчают транспорт воды и растворенных веществ от клетки к клетке.
Эргастические вещества, кристаллические включения и запасные питательные вещества и их локализация в клетках. В процессе жизнедеятельности протопласта возникают разнообразные вещества, получившие обобщенное название эргастических веществ. Они образуются непосредственно в протопласте и отчасти сохраняются в нем в растворенном виде либо в форме включений. В значительно больших количествах эргастические вещества концентрируются вне протопласта, образуя клеточную стенку. Другая часть накапливается в клеточном соке вакуоли в виде растворов или откладывается в цитоплазме в виде разного рода включений. Природа и основные функции эргастических веществ различны. Главнейшие из этих веществ: простые белки, некоторые углеводы, в частности глюкоза, сахароза и крахмал или близкий к нему инулин, а также целлюлоза, запасные жиры и жироподобные вещества — соединения первичного метаболизма; продукты вторичного метаболизма — танниды, полифенольные соединения, алкалоиды, изо пренпро из водные и др. Вещества вторичного метаболизма нередко химически связаны с монои олигосахарами, образуя так называемые гликозилированные формы, или гликозиды. Гликозиды отличаются от негликозилированных соединений прежде всего повышенной способностью проникать через биологические мембраны. К эргастическим веществам относится также обычный во многих растениях оксалат кальция.
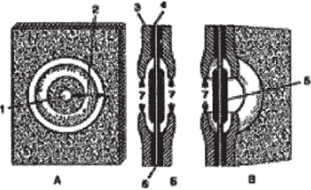
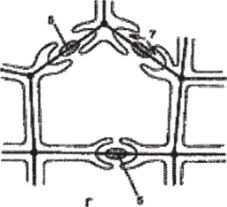
Рис. 89. Строение окаймленных пор в трахеидах хвойных:
А — схема поры в плане; Б — поперечный срез; В — объемное изображение;
Г — поперечный срез трахеиды сосны с окаймленными порами;
- 1 — внутреннее отверстие поры, упирающееся в полость клетки;
- 2 — наружное отверстие поры, упирающееся в первичную оболочку;
- 3 — вторичная оболочка; 4 — первичная оболочка; 5 — торус;
- 6 — срединная пластинка; 7 — внутреннее отверстие поры
Почти все эргастические вещества независимо от природы в той или иной мере могут вновь вовлекаться в процессы активного метаболизма клетки. Поэтому деление этих веществ на ряд групп по их главнейшей функции в известной мере условно.
Важнейшая группа эргастических веществ — запасные вещества. Это белки, углеводы (исключая целлюлозу) и жиры. Главнейшее и наиболее распространенное запасное вещество — полисахарид крахмал. Первичный ассимиляционный крахмал образуется в хлоропластах. Ночью, при прекращении фотосинтеза, крахмал гидролизуется до сахаров и транспортируется в запасающие органы — клубни, луковицы, корневища. Там в особых типах лейкопластов — амилопластах — часть сахаров откладывается в виде зерен вторичного крахмала. Для крахмальных зерен характерна слоистость, что объясняется различным содержанием воды из-за неравномерного поступления крахмала в течение суток. В темных слоях воды больше, чем в светлых. Зерно с одним центром крахмалообразования в центре амилопласта называют простым концентрическим, если центр смещен — простым эксцентрическим. Зерно с несколькими крахмалообразующими центрами — сложное. У полусложных зерен новые слои откладываются вокруг нескольких крахмалообразующих центров, а затем формируются общие слои и покрывают крахмалообразующие центры (рис. 90). Реактивом на крахмал является раствор Люголя, дающий синее окрашивание.
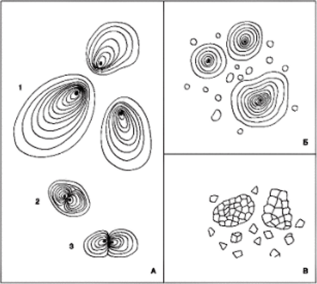
Рис. 90. Крахмальные зерна: картофеля (А):
1 — простое зерно; 2 — полусложное; 3 — сложное; пшеницы (Б), овса (В).
Запасные белки, относящиеся к простым белкам протеинам, чаще откладываются в семенах. Осаждающиеся белки в вакуолях образуют зерна округлой или эллиптической формы, называемые алейроновыми. Если алейроновые зерна не имеют заметной внутренней структуры и состоят из аморфного белка, их называют простыми. Если в алейроновых зернах среди аморфного белка встречаются кристаллоподобная структура (кристаллоид) и блестящие бесцветные тельца округлой формы (глобоиды), такие алейроновые зерна называют сложными (рис. 91). Аморфный белок алейронового зерна представлен гомогенным непрозрачным белком желтоватого цвета, набухающим в воде. Кристаллоиды имеют характерную для кристаллов ромбоэдрическую форму, но в отличие от истинных кристаллов составляющий их белок набухает в воде. Глобоиды состоят из кальциево-магниевой соли, содержат фосфор, нерастворимы в воде и не дают реакцию на белки.
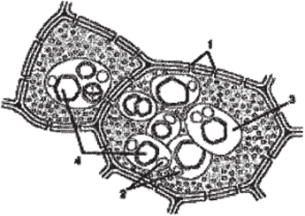
Рис. 91. Сложные алейроновые зерна:
- 1 — поры в оболочке; 2 — глобоиды; 3 — аморфная белковая масса;
- 4 — кристаллоиды, погруженные в амфорную белковую массу
Запасные липиды обычно располагаются в гиалоплазме в виде капель и встречаются почти во всех растительных клетках. Это основной тип запасных питательных веществ большинства растений: наиболее богаты ими семена и плоды. Жиры (липиды) — наиболее калорийное запасное вещество. Реактивом на жироподобные вещества является Судан ГП, окрашивающий их в оранжевый цвет.
Целлюлоза и близкие к ней по химической природе гемицеллюлозы и пектиновые вещества — структурные компоненты клеточной стенки. Продукты вторичного метаболизма выполняют различные функции, но главнейшая из них связана с «организацией» защиты от поедания животными, внедрения болезнетворных микроорганизмов и т. д. Таким образом, они являются группой защитных веществ.
Оксалат кальция чаще всего рассматривается в качестве конечного продукта метаболизма — формы «захоронения» ненужных или вредных клетке веществ. Растения в отличие от животных не имеют специальных выделительных органов и нередко накапливают конечные продукты жизнедеятельности протопласта в виде солей оксалата или карбоната кальция. Кристаллические включения такого типа в значительных количествах накапливаются в тканях и органах, которые растения периодически сбрасывают (листья, корка). Они откладываются исключительно в вакуолях. Форма этих включений достаточна разнообразна: одиночные многогранники, стилоиды (палочковидные кристаллы), пучки игольчатых кристаллов — рафиды, скопления множества мелких кристаллов — «кристаллический песок», сростки кристаллов — друзы (рис. 92). Форма кристаллов нередко специфична для определенных таксонов и иногда используется для их микродиагностики, так для однодольных растений характерны: одиночные призматические кристаллы, стилоиды, рафиды, а для двудольных — друзы, «кристаллический песок», кристаллоносные обкладки.
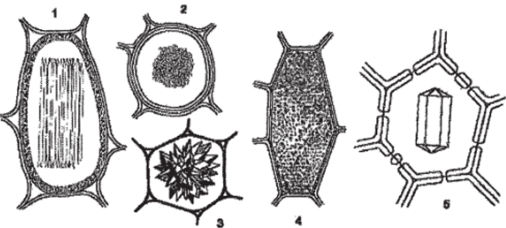
Рис. 92. Формы кристаллов оксалата кальция в клетках:
- 1,2- рафида (недотрога; I вид сбоку, 2 — на поперечном срезе); 3 — друза (опунция); 4 — кристаллический песок (картофель);
- 5 — одиночный кристалл (ваниль)
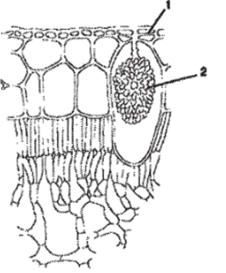
Рис. 93. Цистолит (на поперечном срезе листа фикуса): 1 — кожица листа; 2 — цистолит К кристаллическим включениям близки цистолиты. Они чаще всего состоят из карбоната кальция или кремнезема и представляют собой гроздевидные образования, возникающие на выступах клеточной стенки, вдающейся внутрь клетки. Цистолиты характерны для растений семейств крапивных, тутовых и др. (рис. 93). Возможно, кристаллы и цистолиты выполняют также и защитную функцию, уменьшая шансы поедания этих растений животными.
Обобщенную классификацию эргастических веществ можно представить в виде схемы (рис. 94).
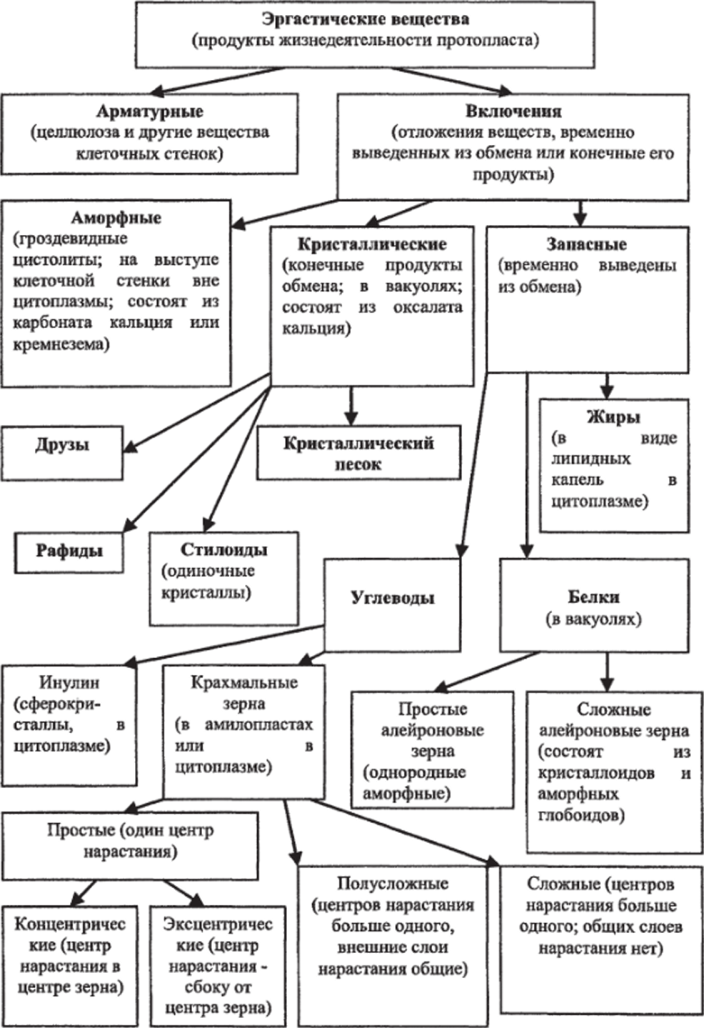
Рис. 94. Классификация эргастнческих веществ.
Тестовые задания для самоконтроля по теме «Особенности строения растительной клетки (цитология)»
1. Назовите пластиды, окрашенные в зеленый цвет:
A) Хлоропласты Б) Лейкопласты.
B) Хромопласты.
2. У однодольных растений кристаллические включения встречаются в виде:
A) Рафид Б) Крахмальных зерен.
B) Алейроновых зерен.
3. Укажите реактив для обнаружения крахмала:
A) Глицерин Б) Кислота уксусная.
B) Реактив Люгсшя.
4. Назовите органоиды растительной клетки, в которых происходит накопление зерен вторичного крахмала:
A) Протеинопласты Б) Амилопласты.
B) Элайопласты.
3. Назовите органоиды растительной клетки, которые определяют цвет листьев осенью:
A) Вакуоли Б) Митохондрии.
B) Хромопласты.
6. У однодольных растений кристаллические включения встречаются в виде:
A) Одиночных призматических кристаллов Б) Друз.
B) Крахмальных зерен.
7. Реактив на лигнин — это:
A) Раствор алюминия хлорида Б) Раствор карбоната натрия.
B) Раствор флороглюцина в НС1 конц.