Обзор литературы.
Изучение влияния добавок различных доз солуната на обмен веществ и энергию роста нетелей

Опытами А. С. Солуна (1956) установлено, что добавка к основному рациону овец витамина, А и Д устраняет яловость животных и повышает их плодовитость на 33%. Автор также сообщает, что процессы воспроизводства у сельскохозяйственных животных часто нарушаются и под комплексным воздействием недостатка двух или нескольких витаминов, поступающих в малых количествах с кормами невысокого качества… Читать ещё >
Обзор литературы. Изучение влияния добавок различных доз солуната на обмен веществ и энергию роста нетелей (реферат, курсовая, диплом, контрольная)
Влияние факторов кормления беременных животных на эмбриональное и постэмбриональное развитие приплода
Продуктивность сельскохозяйственных животных во многом определяется утробным развитием. В этот период происходит закладка и дифферинцировка всех систем и органов, формирование обмена веществ и будущей продуктивности. В литературе имеется достаточно сведений о закономерностях роста и развития животных в эмбриональный и особенно в постэмбриональный периоды (А.А. Малигонов, 1968; С. А. Лапшин, 1972; 1979; 1988; Е. А. Богданов, 1977; Г. А. Богданов, 1981; В. А. Кокорев, 1985; 1986; 1990). Согласно этих и других авторов, наибольшей реакцией на питание обладают интенсивно растущие части. По данным авторов, если в это время усилить питание, то их рост ускорится, и наоборот, если сократить питание, то это оказывает отрицательное влияние прежде всего на них.
Как считают К. Б. Свечин (1961), И. С. Попов (1966), В. И. Шапкарин (1968), В. А. Кокорев (1990) знание такой закономерности дает возможность направленно регулировать рост зародыша на всех этапах эмбриогенеза через питание матери.
Как считает В. А. Кокорев (1990) для того, чтобы побудить зарождающийся организм развиваться в желательном направлении, нужно разработать эффективные методы воздействия, которые должны исходить из глубокого познания биологических процессов, протекающих между матерью и плодом.
Согласно сообщениям Г. А. Шмидта (1952), С. Н. Боголюбского (1968), С. А. Лапшина (1972,1979,1988) условия кормления беременных животных оказывают прямо пропорциональное действие на натальное развитие и рост приплода после рождения. Эти и другие авторы также указывают на то, что на ранних стадиях развития плод хотя имеет очень маленькую массу, правильное, полноценное кормление матери оказывает сильное влияние на их развитие и последующую их продуктивность.
На основании собственных исследований и литературных данных С. А. Лапшин (1972), Ю. В. Никонов (1972), Г. В. Голубев, А. И. Нетеса (1978), А. М. Гурьянов (1979) пришли к выводу, что при неполноценном кормлении беременных животных, особенно на ранней ее стадии, происходит рассасывание зародыша, что в свою очередь может привести к абортам или же рождению слабого и ненормального потомства. Недостаточное или же избыточное питание беременных животных согласно А. А. Малигонова (1968), С. Н. Боголюбского (1975), С. А. Лапшина (1979), В. А. Кокорева (1990) и других, в первую очередь оказывает влияние на те части тела и органы, которые растут наиболее интенсивно в данный период. По данным этих же авторов, недостаточное питание в утробный период задерживает развитие трубчатых костей, что приводит к рождению менее длинноногого и ускозадого молодняка.
Согласно данным Е. А. Махаева (1977) при кормлении супоросных свиноматок в возрасте до двух лет по сниженным нормам, у них в основном увеличивается масса тела, а у взрослых животных это повышение происходит в основном за счет роста матки с плацентой, плодов, плодных вод, их оболочек и молочных желез.
С.Н. Боголюбский (1968), С. А. Лапшин (1972), В. А. Кокорев (1984) считают, что интенсивность роста и развития плода напрямую зависит от обеспеченности материнского организма питательными веществами.
Согласно Н. Г. Макарцеву (2012) в первую треть степени беременности коров и нетелей, хотя энергетический обмен у них увеличивается на 5%, уровень кормления обычно обеспечивает их потребность в питательных веществах и энергии. Автор также указывает, что в этот период особое внимание должно быть обращено на сбалансированность рационов беременных животных по доступному протеину, минеральным веществам и витаминам.
С.А. Лапшин (1988) также считает, что на рост плодов весьма заметное влияние оказывают условия кормления беременных матерей. Так, по данным автора, повышение общего уровня кормления суягных овцематок в начале их беременности на 30%, против рекомендуемых норм, не оказало существенного влияния на рост плодов, а снижение же уровня кормления на 28%, наоборот, привело к снижению массы 45-суточных плодов на 5,3−7,4%.
Автор также считает, что поддержание высокого уровня кормления овцематок в течение всего периода их беременности способствовало увеличению массы 140-суточных плодов на 14,5%, а низкий уровень кормления, наоборот, снижало массу на 7,2%.
Однако в опытах В. А. Кокорева (1984;1990) снижение общего уровня кормления супоросных свиноматок на 20% до 85-суточного периода супоросности не оказало существенного отрицательного влияния на воспроизводительные функции свиноматок, а также на рост и развитие полученных поросят в подсосный период. Противоположного мнения придерживаются Г. Дуск, J .H.Strain (1970). Они считают, что при недостаточном кормлении супоросных свиноматок плодовитость их снижается.
В исследованиях П. Инглыш, У. Смит, А. Маклин (1981) повышение уровня кормления свиноматок находившихся на ограниченном кормлении за 10 суток до случки на 50−100% приводит к увеличению в 2 раза овулировавших яйцеклеток и получению большего количества поросят.
В связи с тем, что вторую треть беременности нетелей плод растет более интенсивно и увеличивает свою массу до 6−8 кг; потребность нетелей в энергии необходимо увеличивать на 14−16% (Н.Г. Макарцев, 2012).
Как считают В. М. Крылов, А. В. Сосновская (1984) в связи с тем, что до 6-ти месячной стельности нетелей развитие плода идет медленно, нетелей в этот период достаточно кормить по нормам для телок старше 14−15 месяцев, а уже на 7−8-ом месяце стельности авторы рекомендуют увеличивать нормы их кормления и они должны соответствовать примерно количеству питательных веществ, необходимых для образования 3−5 кг молока.
Согласно С. А. Лапшину (1988) данные по росту плодов в разные периоды внутриутробного развития при различных уровнях кормления говорят о том, что существующие нормы кормления суягных овцематок обеспечивают нормальное утробное развитие приплода в начале и конце, несколько сдерживают рост плода в середине беременности.
Исследованиями А. И. Нетеси (1977), Г. В. Голубева, А. И. Нетеси (1978) установлено, что по мере повышения норм кормления прирост массы тела свиноматок за период супоросности возрастает, и это в определенной степени влияет и на массу новорожденных поросят, которая, в свою очередь, оказывает существенное влияние на их постнатальный рост и развитие. Таким образом, авторы приходят к выводу о том, что уровень кормления в период супоросности оказывает прямое влияние на массу поросят при отъеме.
Х.Паллсон (1959) считает, что при низком уровне кормления овцематок в первые три месяца суягности, развитие плода происходит нормально, тогда как, в последние месяцы беременности происходит замедление его развития. Однако исследованиями С. Н. Боголюбского (1975) было установлено, что введение в состав рациона овцематок добавочного количества концентрированных кормов в последний месяц их суягности способствует более лучшему развитию эмбриона и рождению крупных и здоровых ягнят. Авторы указывают, что при эмбрионе развивающемся в худших условиях утробного периода, при рождении ягнята были не только мелковесными, но и отличались худшим развитием тела по всем промерам.
В исследованиях С. А. Лапшина (1988) тип кормления суягных овцематок не оказал существенного влияния на развитие плода. Так, плоды овцематок получавших в течение всего периода беременности сенной, силосно-сенной, сенажный и соломо-зерновой рацион одинаковый по питательности существенно не отличались по своему развитию.
В исследованиях Г. А. Хлыстуна (1970), А. А. Хренова (1979), М. А. Наумова (1970; 1989), И. Арнаутовского, Г. Жукова (1981) при кормлении нетелей рационами в структуре которых силос составил от 35 до 55% по питательности, получали более жизнеспособных телят.
Развитие плода и его жизнеспособность обусловлены характером и условием обмена веществ в организме матери, степенью обеспеченности и использования питательных веществ рациона (В.А. Кокорев, 1990) .
Существенное влияние на развитие плода оказывает содержание протеина в рационе. Так, в серии опытов проведенных С. А. Лапшиным (1972; 1988) по изучению влияния уровня протеина в рационах овцематок в разные периоды беременности на рост плода, в котором для контроля была взята группа овец с умеренным уровнем протеина — по 112 г на голову в сутки. Три группы получали по 169 г протеина (высокий уровень), одна из которых это количество протеина получала в течение всего периода беременности, другая в первую, третья во вторую половину беременности. Другие три группы получали по 85 г (низкий уровень) протеина, при этом одна группа получала это количество протеина в течении всей беременности, вторая в первую половину, третья во вторую половину беременности. В результате этих опытов автором было установлено, что уровень протеина не оказывает существенного влияния на рост тела эмбриона в первую треть беременности.
Повышение или снижение уровня протеина в рационах овцематок стало сказываться на росте плодов только к 75-дневному возрасту. При повышенном уровне протеина масса плодов в этом возрасте увеличилась на 7,2−7,7%, а при пониженном — уменьшилась на 8,4−8,6%. В конце утробного периода разница в массе плодов при разных уровнях протеина в рационах несколько увеличилась. Так, в группе высокого уровня протеина в течение всей беременности масса 140-дневных плодов стала выше контрольных на 13,2%, в группе высокого уровня во вторую половину беременности на 6,6%. При низком уровне протеина в течение всей беременности масса плодов была ниже, а 10,8%, а во вторую ее половину на 9,5%. Повышение уровня протеина в первую половину беременности не оказало существенного влияния на дальнейший рост плодов, а отставание в росте плодов при его снижении в это время полностью не компенсируются до родов даже при улучшенных условиях кормления матерей.
Положительные результаты при повышении количества протеина в рационах беременных животных были получены и в опытах И. Г. Елисеева, Б. Н. Брандиса (1962), А. И. Петрова, А. В. Метлицкого, А. И. Ибраева (1982). Так, этими авторами было установлено, что повышение уровня протеина в рационах беременных животных по сравнению с принятыми нормами на 20−30% увеличивает плодовитость на 30% и способствует рождению крупного и жизнеспособного приплода. Аналогичные результаты на супоросных свиноматках были получены А. В. Квасницким (1959), на овцематках S.B.Slen (1953), E.W.Klesterman, D.W.Bolin, M.Z.Buchawan, F.M.Bolin, W.E.Denissen (1953).
Согласно данных Aghina Cesare (1978) чрезмерное кормление коров в течение 40 дней высоко белковым кормом, превышающее потребность на 5−6%, вызывает нарушение овуляции и гибель эмбриона уже на ранних стадиях.
Положительное влияние на развитие плода оказывают использование в рационах беременных животных синтетических заменителей кормового протеина.
Так, в опытах С. А. Лапшина (1988) скармливание карбамида в умеренных дозах не оказало отрицательного влияние на состояние суягных овцематок и на утробный рост ягнят. В частности, скармливание овцематкам по 10 г карбамида в сутки дополнительно к основному рациону, недостаточному по переваренному протеину, не оказало заметного влияния на рост эмбрионов в первую треть беременности, но способствовало увеличению массы плодов во вторую треть — на 2,7% и последнюю треть беременности — на 6,9%.
Известно, что биохимические функции у плода сельскохозяйственных животных проявляются уже с первых дней эмбрионального развития. В этот период начинают формироваться многие породные и хозяйственно-полезные признаки. Поэтому, данные обмена веществ на разных стадиях внутриутробного развития плода помогают понять факторы, регулирующие процессы формообразования плода (Д.Ш. Гайирбегов, 2002).
Существенное влияние на внутриутробное развитие приплода оказывают минеральные вещества. Известно, что потребность в них во многом определяется физиологическим состоянием организма. Она особенно велика у растущих животных, во время беременности и лактации. У беременных животных наблюдаются повышение обмена веществ, активация процессов ассимиляции, создание запасов элементов питания, исключительно бурный рост матки с плацентой и плода на основе ресурсов материнского организма (В.А. Кокорев, 1988).
В литературе имеются достаточно сведений указывающие о важной роли минеральных веществ в рационах беременных животных на формирование плода (С.А. Лапшин, 1972, 1988; Ю. В. Никонов, 1973; А. М. Гурьянов, 1979; В. П. Помпаев, 1982; В. П. Ходыков, 1982; Д. Ш. Гайирбегов, 1986, 2002; В. В. Демин, 2004; Ц. Б. Тюрбеев, 2005; Ц. О. Улюмджиев, 2009) и другие.
Как считает С. А. Лапшин (1988), наибольшее влияние на внутриутробное развитие плода оказывают содержание в рационах беременных животных кальция и фосфора. Вместе с тем, автор в своих исследованиях установил, что повышение уровня кальция и фосфора в рационах беременных овцематок на 28 и 56% сверх рекомендуемых норм за счет скармливания костной муки не оказывает существенного влияния на массу плода в течение всего утробного периода развития.
С.А. Лапшин (1988) также считает, что при наличии в рационах суягных овцематок фосфора в количестве 7,2 и 9,8 наблюдается лишь тенденция к увеличению массы плода к концу беременности. Согласно этого же автора, недостаток этих двух макроэлементов приводит к нарушению развития плода, и особенно его скелета, к рождению слабых, пониженной жизнеспособностью ягнят. Восполнение же недостатка данных элементов питания за счет минеральных подкормок ликвидирует отрицательные явления в развитии плода, а также усиливает рост периферического скелета и способствует повышению массы ягнят при рождении.
Б.С. Орлинский (1984) также считает, что использование для супоросных свиноматок рационов с пониженным на 25% уровнем кальция и фосфора снижает плодовитость, кроме того проявляются аномалии в развитии эмбрионов, рождается слабое потомство, увеличивается смертность поросят при рождении. Об аналогичных проявлениях указывают также В. А. Гуляев (1979), В. А. Кокорев (1984), H.R.Dawigson (1930) и другие.
Также, в опытах В. А. Кокорева (1984;1990) самое высокое многоплодие было у свиноматок, которые, на фоне полноценного кормления, получали оптимальное количество кальция (30 г на голову в сутки). Снижение или повышение этого уровня на 30% приводило к достоверному уменьшению многоплодия. На основании анализа полученных в эксперименте данных автор приходит к заключению, что оптимизация уровня кальция в рационах свиноматок способствует увеличению плодовитости, получению более крупных поросят, которые лучше росли и развивались в подсосный период. Аналогичное положительное действие на репродуктивные функции свиноматок, по данным этого же автора, оказало и оптимизация количество фосфора в их рационах.
Как сообщает С. А. Лапшин (1988), увеличение содержания фосфора в рационах суягных овцематок также не оказало существенного влияния на рост плодов. Наблюдалась лишь тенденция к увеличению массы плодов только в конце беременности маток, при наличии в их рационах от 7,2 до 10,8 граммов фосфора.
На воспроизводительные функции животных, рост и развитие плода значительное влияние оказывают содержание в рационах беременных животных и других макроэлементов. Так в опытах, проведенных Н. Б. Тюрбеевым (2005), на суягных овцематках установлено, что оптимизация в их рационах количества калия и натрия создает благоприятные условия для внутриутробного плода, увеличивает многоплодие и сохранность ягнят. Автор также указывает, что различные уровни натрия и калия в рационах овцематок оказали влияние на онтогенез плода в течение всего периода суягности маток. Так, к концу утробного развития эмбрионы матерей, которые получали оптимальный уровень этих элиментов имели преимущество по сравнению с эмбрионами от матерей получавших повышенное и пониженное количество калия и натрия в ширине груди на 13,4 — 15,7%, глубине груди на 3,4−8,0%, по обхвату груди на 1,4 — 5,3%.
Об увеличении многоплодия, в частности у супоросных свиноматок, под действием оптимального количества калия и натрия в рационе сообщает также В. А. Кокорев (1990). Увеличение же количества калия в рационах супоросных свиноматок, наоборот, приводит к снижению многоплодия и крупноплодности (В.А. Кокорев, А. М. Гурьянов, 1980).
Согласно Э. Визнера (1976) при обильном потреблении калия плодовитость самок понижается уже во втором поколении. Автор также указывает, что недостаток натрия вызывает нарушение охоты и бесплодия у овец.
Как сообщает В. А. Кокорев (1990), изменение уровня магния в рационах также оказывает своё влияние на продуктивность свиноматок. Так, по данным автора, самое высокое многоплодие (13 гол) отмечалось у маток, получавших оптимальную норму магния (7.3 г на голову в сутки). Пониженный его уровень (5.2 г) приводил к снижению многоплодия свиноматок на 17.7%, а повышенный уровень элемента также давал подобный же эффект — уменьшение этого показателя на 12,3%. Однако, как указывает Э. Визнер (1976), при учете значения магния для животных этот элемент в смысле управления воспроизводством вряд ли заслуживает особого признания.
Среди минеральных веществ большое значение для нормального размножения, роста и развития плода имеют и микроэлементы. Об этом сообщают Ю. В. Никонов (1973), С. А. Лапшин (1988), П. М. Помпаев (1982), В. В. Демин (2007), Ц. О. Улюмджиев (2009) и другие.
Так, в опытах Ю. В. Никонова (1973), подкормки суягных овцематок молибденовокислым аммонием в дозе 5 и 10 мг на голову в сутки несколько сдерживает рост эмбрионов во время зародышевого и предплодного периодов на — 5,4%, не оказывает отрицательного влияния в середине и заметно улучшает их рост (на 7,7 и 8,4%) в конце плодного периода.
Об аналогичных результатах сообщает также и С. А. Лапшин (1988). По мнению этих авторов, воздействие молибденовокислого аммония в разных дозах на рост плода было примерно одинаковое.
По данным Д. Ш. Гайирбегова (1986; 2002) оптимизация количества молибдена в рационах стельных коров и суягных овцематок, за счет подкормки их молибденовокислым аммонием, способствовала получению более крупного и жизнеспособного приплода. Такие же положительные результаты при использовании в рационах супоросных свиноматок молибденово содержащей добавки были получены и П. М. Помпаевым (1982).
В исследованиях С. А. Лапшина (1988) по изучению влияния разных уровней марганца, кобальта, меди в рационах суягных овцематок на внутриутробный рост приплода было установлено, что скармливание солей этих элементов оказывает довольно существенное влияние на рост плода в течение всего утробного периода. Наиболее же сильно оно проявилось в начале утробного развития и в дальнейшем с возрастом это их влияние снижается. Автор также сообщает, что добавка в рационы овцематок хлористого кобальта в количестве 5 мг на голову в сутки способствует увеличению массы 45-суточных плодов на 27,7%, 90-суточных — на 140% и 130-суточных — на 4%. Повышенная дача соли кобальта, в количестве 10 мг, в начале утробного развития не оказывает положительного действия, а в середине и конце несколько интенсифицирует рост плода по сравнению с меньшей дозировкой (5 мг) хлористого кобальта.
По данным Э. Визнера (1976) из всего многообразия различных функций марганца в обменных процессах вытекает его значение не только для формирования скелета и функционирования мускулатуры, но и непосредственно для развития и деятельности полового аппарата животных обоего пола.
Как сообщает A. Henning (1967) марганец является составной частью ферментов, которые необходимы для синтеза определенных половых гормонов. При недостатке этого элемента у жвачных животных происходят нарушения половой деятельности и они плохо оплодотворяются (Э.Визнер, 1976).
В опытах В. П. Ходыкова (1982) применение в рационах супоросных свиноматок марганца в оптимальной дозировке, способствовало повышению многоплодия и получению жизнеспособных поросят.
Как сообщает С. А. Лапшин (1988) подкормка суягных овцематок сернокислым марганцем оказывает несколько меньшее положительное влияние на рост плода, чем хлористого кобальта. Так его подкормки в количестве 10 мг обеспечили увеличение массы плода в начале утробного развития на 17,2%, в середине — на 14,5%, в конце — на 9%. Его количество в 20 мг, по мнению автора, дает меньший эффект, чем 10 мг, и обеспечивает увеличение массы плода в начале утробного развития — на 15%, в середине — на 10,7% и в конце — 5,7%. С возрастом плода, сообщает автор, эффективность подкормок сернокислым марганцом снижается.
Согласно исследованиям В. А. Кокорева (1971) на процессы воспроизводства и рост плода животных существенное влияние оказывают соли меди и цинка. Так, по данным автора, скармливание в составе рациона 10 мг сернокислой меди суягным овцематкам способствовало увеличению массы плода на 27,2%; длины плода — на 12,2%; обхвата груди — на 4,9%; 25 мг сернокислой меди — соответственно на 33,9%, 11,4% и на 18,7%. Совместное же скармливание 10 мг сернокислой меди и 5 мг сернокислого цинка, по данным этого же автора, существенно не изменяло рост плодов.
Автор также сообщает, что подкормки сернокислой меди оказали положительное достоверное влияние на рост сердца, печени, легких, почек и желудка в начале и в конце, а добавки сернокислого цинка на органы пищеварения в конце внутриутробного периода.
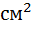
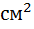
В опытах С. С. Маштыкова (2011) добавка в рационы беременных овцематок каракульской породы кремний и железо содержащей кормовой добавки «Феросил» в оптимальной дозе (5 мг/кг живой массы) способствовало лучшему росту матки с плацентой, увеличению котиледонного аппарата в начале беременности на 6,8, в середине — на 18,8 и в конце — на 33,0, количества крови, протекающей через средние маточные артерии — на 2,67%, а также лучшему развитию плода в течение всего эмбрионального периода, по сравнению с группой маток не получавшими кормовую добавку. Автор также отмечает, что кремний и железосодержащая добавка способствовала рождению крепких и жизнеспособных ягнят, обеспечивает дальнейший стабильный и интенсивный их рост.
На четко установленное значение йода для воспроизводства у супоросных свиноматок указывают Г. Г Смирнов (2000), Е. В. Громова, С. Г. Кузнецов (2003). По данным авторов, оптимизация уровня этого микроэлемента в рационах свиноматок в 2/3 и 1/3 половины супоросности способствовала увеличению многоплодия, лучшему росту и развитию поросят в подсосный период.
На аналогичное положительное действие по увеличеню плодовитости — на 4%, крупноплодности — на 7,8%, массы гнезда при рождении — на 12,1% и массы гнезда в месячном возрасте — на 22,2% при восполнении дефицита минеральных элементов питания в рационе супоросных свиноматок указывает также и С. В. Максимов (1999).
Ц.О. Улюмджиев (2009) изучая влияние добавок в рационы суягных курдючных овцематок разных уровней селен содержащего препарата ДАФС-25 на внутриутробное развитие ягнят установил, что эмбрионы от матерей которые получали добавки препарата в оптимальных количествах имели преимущество по сравнению с контрольными, как по массе, так и по всем промерам во все периоды беременности овцематок. Так, в начале утробного развития по массе они превосходили контрольных на 16,8%, по длине туловища — на 11,3%, по ширине груди — на 8,4%, глубине груди — на 24,7% и обхвату груди за лопатками — на 10,2%. Аналогичная же картина по массе и промерам наблюдалось в середине и в конце утробного развития плодов.
На рост и развитие приплода существенное влияние оказывает наличие в рационе беременных животных и витаминов. Многочисленными исследованиями установлено, что витамины способствуют воспроизводству только тогда, когда они поступают в определенной пропорции друг к другу (Э.Визнер, 1976).
Как сообщает автор, правило, гласящее, что одностороннее введение витаминов вредно, подтверждается в отношении воспроизводства для многих витаминов, но особенно в отношении витамина А.
У морских свинок из-за избыточного потребления аскорбиновой кислоты значительно понижалась плодовитость. Если животным ввести в избытке витамин Д, то это вызывает повышенную раздражительность матки, что может вызвать аборт.
Опытами А. С. Солуна (1956) установлено, что добавка к основному рациону овец витамина, А и Д устраняет яловость животных и повышает их плодовитость на 33%. Автор также сообщает, что процессы воспроизводства у сельскохозяйственных животных часто нарушаются и под комплексным воздействием недостатка двух или нескольких витаминов, поступающих в малых количествах с кормами невысокого качества. В своих исследованиях H. Konermann (1967) убедительно демонстрирует связь воспроизводства у крупного рогатого скота с обеспечением их каротином. Выведенный им коэффициент регрессии +0.1 показывает, что с увеличением дефицита каротина до 100 мг на одну голову в день интервал между первым осеменением и успешным оплодотворением превысил 10 дней. Исследованиями английских ученых S. Metrel, H. Ritter (1955) установлено, что витамин, А необходим, как для овуляции и оплодотворения, так и для имплантации зиготы.
Согласно данным K.J.Dawey, J.W.Stewensona (1963) в опытах, проведенных на нескольких поколениях свиней, где использовались рационы с содержанием 7, 11, 14 и 17 мг пантотеновой кислоты, было установлено, что в группе, получавшей корм с самым низким содержанием этого витамина, величина помета отчетливо уменьшалась уже во втором поколении.
Как указывает Г. П. Белехов, А. А. Чубинская (1960) при недостатке в рационах свиноматок рибафловина, поросята рождаются крайне слабые или же мертвые. Добавка же данного витамина в количестве 2,7 мг на 1 кг сухого вещества корма полностью восстанавливает нормальное размножение и рождение поросят.
Таким образом, анализ существующих отечественных и зарубежных литературных данных показывает, что заботу о получении крепкого жизнеспособного приплода следует проявлять уже в период их эмбрионального развития, когда происходит основная закладка, дифференцирование всех систем и органов, а также формирование будущей продуктивности, которые, в свою очередь, во многом определяются полноценным питанием материнского организма.