Свойства Na+-и Крюков, вызывающих генерацию потенциала действия

Другими специфическими свойствами натриевого тока являются его нелинейная зависимость от деполяризации мембраны и способность вызывать два разных по кинетике ответа — небольшой, сравнительно медленный допороговый ответ (локальный) и быстрый ответ в виде резко нарастающей самоусиливающейся деполяризации мембраны, возникающий при сверхпороговой деполяризации. Зависимость процесса активации… Читать ещё >
Свойства Na+-и Крюков, вызывающих генерацию потенциала действия (реферат, курсовая, диплом, контрольная)
Натриевый ток во время генерации ПД обладает рядом особенностей. Первой особенностью является его быстрая активация. Действительно, при мгновенной деполяризации мембраны до -9 мВ возникающий Ыа+-ток нарастает до своего пика всего за 0,1—0,2 мс. Было показано, что крутизна нарастания натриевого тока является потенциал-зависимой, и чем больше деполяризация мембраны, тем быстрее открываются Ыа+-каналы, а следовательно, быстрее активируется Na+-TOK.
Второй важнейшей особенностью потенциал-зависимого Na ‘ -тока является его быстрое затухание вплоть до исчезновения, происходящее вслед за его активацией и получившее название инактивации №+-тока. Инактивация Na+-TOKa происходит по той же причине, что и его активация, т. е. она вызвана деполяризацией мембраны. При этом инактивация протекает в 10 раз медленней активации. Это позволяет каналу быстро открыться (активироваться), пропустить ток и закрыться (инактивироваться). Ясно, что инактивация ограничивает время протекания тока по Ка+-каналам. В отсутствие инактивации Ыа^-каналы могли бы оставаться открытыми, а натриевые токи — продолжать течь, что значительно затруднило бы развитие второй фазы ПД — реполяризации.
Способность №+-каналов и Ыа+-тока к инактивации имеет большой биологический смысл и обусловливает чрезвычайно важное свойство возбудимой мембраны — ее рефрактерность.
Рефрактерностъ — это состояние невозбудимости мембраны, или временная неспособность мембраны к возбуждению и генерации ПД. Наблюдения за состоянием возбудимости мембраны показали, что в момент генерации ПД и сразу же после него нельзя вызвать вновь генерацию IIД при использовании любых сил раздражающего тока. Этот интервал, соответствующий полной невозбудимое™ мембраны, получил название абсолютного рефрактерного периода, или абсолютной рефрактерности. У гигантского аксона кальмара период абсолютной рефрактерное™ составляет около 1 мс, у других объектов этот период может составлять десятки и сотни миллисекунд. Причина рефрактерное™ кроется в том, что во время первой фазы ПД практически все Иа^-каналы в месте деполяризации мембраны уже активированы и нет незадействованиых в активации каналов. К моменту развития пика ПД практически все Иа+-каналы инактивированы, а значит, не могут быть вовлечены в генерацию нового ПД. На рис. 2.7, а видно, что состояние абсолютной рефрактерное™ длится на протяжении всей фазы деполяризации и начальной стадии фазы реполяризации.
Вслед за периодом абсолютной рефрактерное™ следует интервал, когда потенциал действия можно вызвать повторно. Однако чем меньше прошло времени после окончания периода абсолютной рефрактерности, тем меньше будет амплитуда повторного потенциала действия и тем большая сила нужна, чтобы его вызвать. В этот период, получивший название.
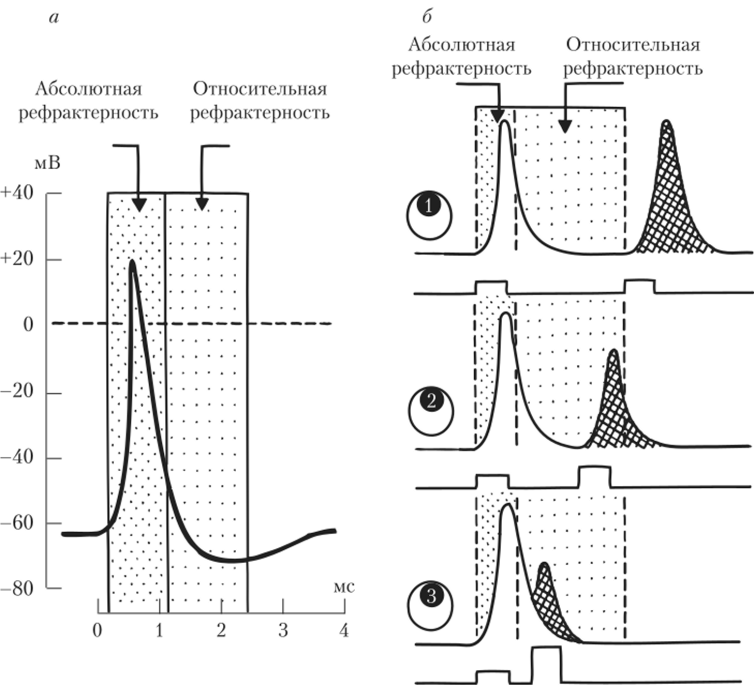
Рис. 2.7. Периоды абсолютной и относительной рефрактерности (невозбудимое™) мембраны при генерации ПД (а) и ответные реакции мембраны при генерации второго ПД (б):
О — последующий ПД не отличается от первого, вызывается таким же, как и первый, пороговым стимулом; © и (c) — мембрана пребывает в состоянии относительной рефрактерности (ПД, следующий за первым, вызывается сверхпороговым стимулом и имеет сниженную амплитуду) периода относительной рефрактерности, возбудимость мембраны клетки понижена, а порог повышен. Однако постепенно возбудимость мембраны достигает исходной величины, а амплитуда повторного потенциала действия достигает нормы (рис. 2.7, в). Следует отметить, что период относительной рефрактерности, занимающей в аксоне кальмара 1,5 мс, варьирует в довольно широких пределах от объекта к объекту.
Рефрактерность создает ограничения для ритмической работы возбудимых клеток, выполняя роль своеобразного частотного фильтра. Этот механизм позволяет предотвратить утомление нервных клеток. Например, при продолжительности периода абсолютной рефрактерности 4 мс максимальная «пропускная» частота нервного волокна равна 250 Гц.
С явлением рефрактерности тесно связано понятие о лабильности (функциональной подвижности) возбудимых тканей, введенное русским физиологом Николаем Евгеньевичем Введенским. Мерой лабильности является количество потенциалов действия, которое способна генерировать возбудимая ткань в единицу времени. Очевидно, что лабильность возбудимой ткани в первую очередь определяется продолжительностью периода рефрактерности. Наиболее лабильными являются волокна слухового нерва, в которых частота генерации потенциалов действия достигает 1000 Гц.
Ограничение частоты возбуждения рефрактерностыо является решающим фактором для нормального функционирования не только аксонов, но и кардиомиоцитов, как системы, обеспечивающей насосную функцию сердца. Длительная рефрактерность клеток миокарда обеспечивает паузы в несколько миллисекунд, необходимые для заполнения сердца новой порцией крови после очередного сокращения, а также препятствует неконтролируемой циркуляции возбуждения по миокарду.
Другими специфическими свойствами натриевого тока являются его нелинейная зависимость от деполяризации мембраны и способность вызывать два разных по кинетике ответа — небольшой, сравнительно медленный допороговый ответ (локальный) и быстрый ответ в виде резко нарастающей самоусиливающейся деполяризации мембраны, возникающий при сверхпороговой деполяризации. Зависимость процесса активации Ыа+-канала от деполяризации мембраны имеет S-образную форму (рис. 2.8). При незначительных деполяризационных сдвигах мембранного потенциала степень активации Na+-каналов относительно невелика. Поэтому в ответ на небольшую донороговую деполяризацию мембраны наряду с пассивным ответом на мембране возникает лишь небольшой активный локальный ответ, связанный с открытием небольшого числа Na^-каналов и развитием небольшого входящего натриевого тока.
Локальный ответ пропорционален вызывающей его силе, поэтому он растет по мере возрастания деполяризации мембраны (и натриевого тока). Однако в ответ на деполяризацию активируются не только натриевые, но и небольшое число потенциал-зависимых К+— каналов. Поскольку К+-ток имеет более медленный ход, то локальный ответ успевает развиться за счет входа Na+, но постепенно гасится противоположно направленным К^-током (/Кг) в сочетании с К+-током утечки (/к утечки)* При критическом уровне деполяризации мембраны наступает момент, когда скорость нарастания входящего Ыа+-тока и его величина /Na становится больше (или равной) сумме двух гиперполяризующих калиевых токов — потенциал-активируемого калиевого тока (/Кг) и калиевого тока утечки (/к утечкн), т. е. /Na > IKv + /к ки. В результате возрастающий Ыа+-ток уже не компенсируется более медленно развивающимися выходящими гиперполяризующими калиевыми токами (IKz) + /к ухсчки). С этого момента — резкого возрастания скорости входящего Na^-тока — и начинается процесс генерации собственно ПД (рис. 2.8, а).
Для оценки числа Na^-каналов, переходящих в открытое состояние в зависимости от деполяризации мембраны, служит коэффициент т, который описывает долю Ыа+-каналов от их общего числа, которые активируется при данном мембранном потенциале. На рис. 2.8, б показана зависимость.
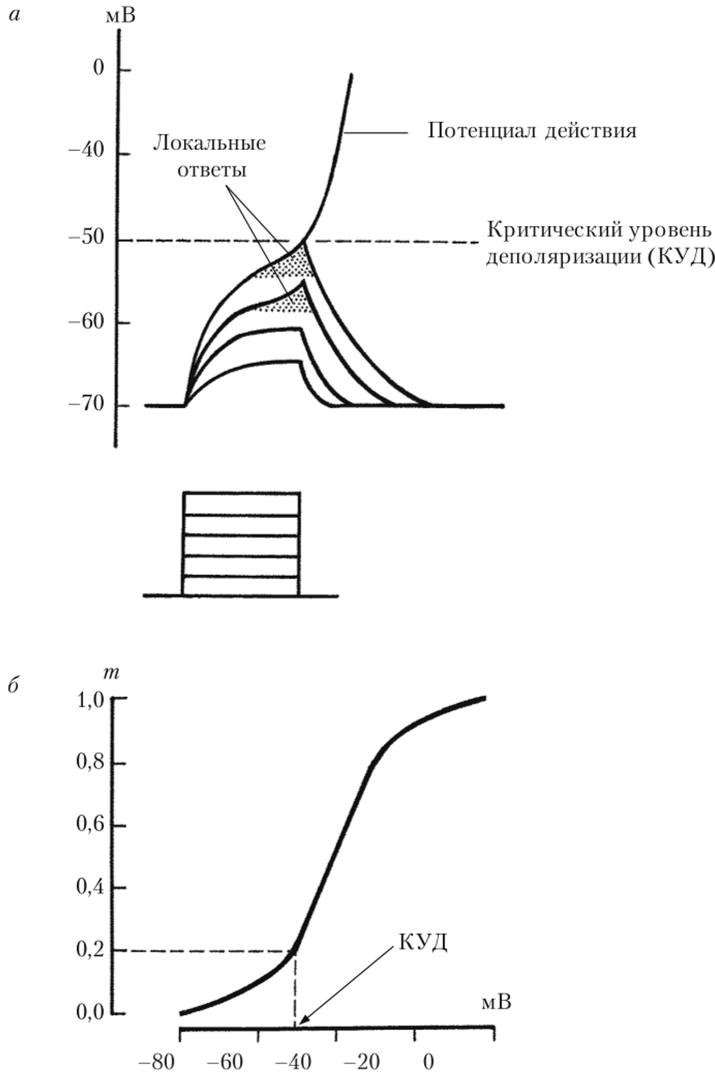
Рис. 2.8. Пассивные сдвиги мембранного потенциала клетки в ответ на деполяризующие токи, пропускаемые через мембрану с помощью внутриклеточного электрода (я), и потенциал-зависимость коэффициента т (б) (показывает, какова доля №+-каналов с открывшейся ш-створкой при данном мембранном потенциале) значения коэффициента т от потенциала на мембране. При потенциале покоя все Ыа+-каналы закрыты и т = 0. При небольшой деполяризации мембраны (вплоть до -50 мВ) кривая потенциал-зависимости коэффициента т достаточно пологая. Следовательно, Ыа+-ток при небольших допороговых деполяризациях невелик (т = 0,1 0,2) и медленно нарастает. При сверхпороговых деполяризациях (выше КУД) крутизна кривой потенциал-зависимости коэффициента ш значительно возрастает. Это значит, что быстрее нарастает число активированных Ка+-каналов и, следовательно, величина Ыа+-тока. Это приводит к появлению самоусиливающегося регенеративного Ыа+-тока.
На рис. 2.9 показан цикл Ходжкина, который демонстрирует, как деполяризация до уровня КУД, приводя к открытию небольшого дополнительного числа Ыа+-каналов, вызывает дополнительную деполяризацию. Она, в свою очередь, стимулирует открытие новых Ыа+-каналов, а значит, и новую, дополнительную деполяризацию мембраны входящим натриевым током. Возникает эффект самоусиления натриевого тока за счет создаваемой им деполяризации мембраны. Наличие такой положительной обратной связи между Na^-током и деполяризацией является важным условием для необратимого самозапуска первой деноляризационной фазы IIД.
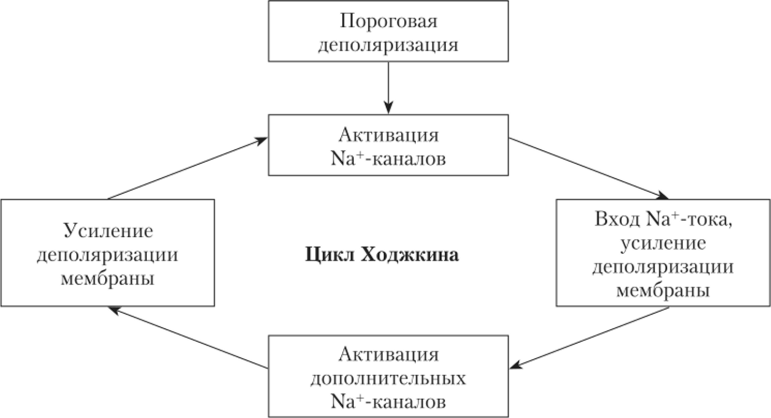
Рис. 2.9. Схема цикла Ходжкина, демонстрирующая положительную обратную связь между деполяризацией мембраны и входящим в клетку Na+-TOKOM, усиливающим и ускоряющим деполяризацию.
Калиевый ток при генерации ПД также отличается рядом особенностей. Во-первых, он развивается в 10—15 раз медленней, чем натриевый. В ответ на мгновенную деполяризацию мембраны, вызванную током натрия, калиевый ток, начиная нарастать практически одновременно с натриевым, достигает максимума только через 1 мс, когда Ыа+-ток уже спадает, преодолев пиковое значение. Во-вторых, именно К+-ток во время генерации ПД возвращает изменившийся до положительных величин мембранный потенциал к исходному значению. Он как бы «выпрямляет» возникшее при генерации ПД быстрое отклонение мембранного потенциала от уровня потенциала покоя. Поэтому потенциал-зависимый К+-ток (Ка-ток), текущий на второй фазе ПД, принято называть К+-током задержанного выпрямления (англ, delayed-rectifier).
Еще одной отличительной особенностью Кг;-тока является отсутствие у него инактивации. В результате в течение всего периода деполяризации мембраны этот ток продолжает течь через мембрану.
Специфическим свойством К+-тока является его линейная зависимость от деполяризации мембраны. Величина К+-тока будет непрерывно возрастать по мере роста деполяризации мембраны на первой фазе ПД. Близкая к линейной зависимость величины К+-тока от возрастающей деполяризации мембраны будет обеспечивать отрицательную обратную связь в процессе генерации ПД. По мере роста деполяризации К+-ток будет увеличиваться, противодействуя ей и ускоряя возвращение мембранного потенциала к исходному уровню потенциала покоя.