Теоретические положения хода роста и производительности искусственных древостоев

Массовые обследования искусственных древостоев показывают, что темпы хода роста и накопления стволового запаса не носят плавного характера. Это хорошо иллюстрируют кривые изменения корневого запаса сосновых насаждений в условиях лесной опытной дачи ТСХА (рис. 11.5). На этом рисунке Г. Ф. Михальченко (1989) показала фактически не только динамику возрастания корневого запаса к 45—50 годам, но и его… Читать ещё >
Теоретические положения хода роста и производительности искусственных древостоев (реферат, курсовая, диплом, контрольная)
Известно, что чем больше густота посадки лесных культур, тем они раньше смыкаются, и чем больше густота посадки, тем интенсивнее идет процесс естественного изреживания насаждения. При этом оказываются изменяющимися с возрастом не только сомкнутость, но и полнота, и класс бонитета (Разин, 1967, 1977; Рубцов, 1969).
Насколько интенсивно идет ход естественного изреживания можно судить наглядно по рис. 11.3, где показана возрастная динамика густоты стояния в культурах сосны с разной густотой посадки. Графическое изображение динамики естественного изреживания выполнено на основе многолетних учетов опытных культур, созданных проф. М. К. Турским в 1879 г. на территории Лесной опытной дачи. На протяжении 130-летнего периода роста этих искусственных сосняков в них не проводились рубки промежуточного пользования, благодаря чему можно судить о динамике естественного роста и отпада модельных популяций.
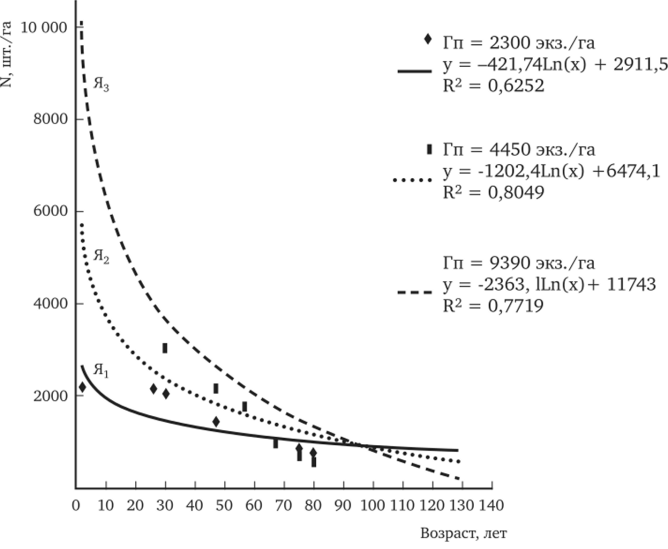
Рис. 11.3. Динамика естественного изреживания сосны в культурах с разной густотой посадки (Гп)
Опытные культуры представлены однолетними сеянцами на трех пробных площадях со следующей густотой посадки, шт./га: 2300 (пр. пл. Я,), 4450 (пр. пл. Я2) и 9390 (пр. пл. Я3) на 1 га. Как видно из рис. 11.3, несмотря на то, что крайние варианты густоты посадки изначально отличались в 4 раза, к возрасту 95 лет численность стволов на 1 га фактически сравнялась на всех трех пробных площадях. Причем культуры сосны с густотой посадки 9390 шт./га по интенсивности отпада превышали почти в 6 раз таковой в культурах с очень редкой густотой посадки. Мало того, в культурах с густотой посадки 9390 шт./ га отпад принял регрессирующий характер. Это привело к тому, что к 130 годам очень редкие смолоду культуры стали наиболее густыми: их густота стояния почти в 3 раза превысила некогда более густую посадку.
Г. С. Разиным (1979) был проведен фундаментальный анализ динамики сомкнутости древостоев различной начальной густоты (рис. 11.4), произрастающих в приблизительно равных лесорастительных условиях. Им выявлено, что древостой, являющиеся наиболее сомкнутыми в молодом возрасте, не будут такими же в старших классах возраста; древостой становятся менее сомкнутыми тем раньше, чем раньше они достигают предельной сомкнутости. Для каждого древостоя имеется определенный естественный порог, после прохождения которого древостой не может быть возвращен рубками разреживания на стадию (линию) прогрессивного развития — стадию повышения сомкнутости крон. Таким порогом является индивидуальная предельная сомкнутость крон. Если древостой естественным изреживанием преодолеет этот порог и перейдет в стадию регресса, то процесс становится необратимым полностью или частично в зависимости от степени деградации насаждения.
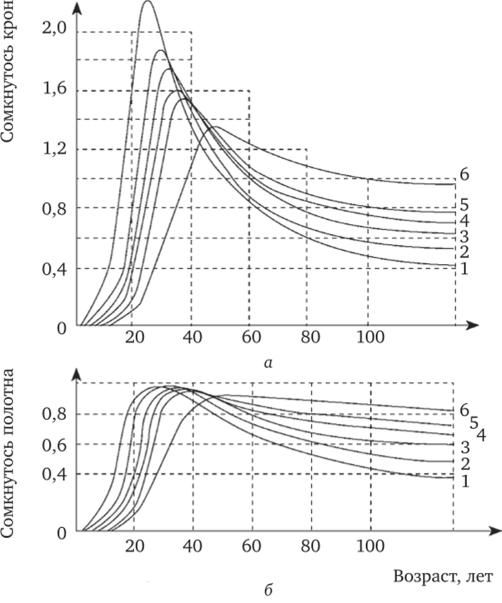
Рис. 11.4. Возрастная динамика сомкнутости крон (а) и сомкнутости полога (б) одноярусных еловых древостоев различной начальной густоты (в возрасте 5 лет), тыс. экз./га, около:
1 — 67 тыс. экз.; 2 — 22 тыс. экз.; 3 — 11,5 тыс. экз.; 4 — 7 тыс. экз.; 5 — 4,5 тыс.
экз.; 6 — 1,8 тыс. экз.
Согласно исследованиям Г. С. Разина (1979), большинство древостоев в естественных условиях оказываются с возрастом недостаточно сомкнутыми, полными и продуктивными. Такими они становятся намного раньше достижения ими возраста технической спелости и главной рубки. Основной причиной, как правило, является большая начальная густота деревьев, приводящая к взаимному угнетению и ослаблению большинства деревьев и всего древостоя в целом. Ошибочно вести речь о естественном ходе роста сомкнутых (нормальных, полных) древостоев, так как древостой при естественном ходе роста не могут, начиная смолоду, оставаться длительное время сомкнутыми. Поэтому таблицы хода роста, составленные для полных деревьев, не могут отражать реальной картины их роста и производительности.
По этой причине, например, составленные А. М. Бородиным (1972), таблицы хода роста полных (нормальных) еловых культур в Московской области по классам бонитета не могут считаться объективными. Так, по его таблицам при 16 классе бонитета в возрасте 80 лет культуры ели имеют запас стволовой древесины 1210 м3/га. Это крайне завышенные данные, ибо нами даже при обследовании самых лучших высокопродуктивных тюрмеровских культур, да и вообще ельников зоны смешанных лесов, столь высокие запасы нигде не наблюдались. Такая продуктивность возможна лишь для насаждений ели европейской в горных условиях Европы (Карпаты, Судеты).
Массовые обследования искусственных древостоев показывают, что темпы хода роста и накопления стволового запаса не носят плавного характера. Это хорошо иллюстрируют кривые изменения корневого запаса сосновых насаждений в условиях лесной опытной дачи ТСХА (рис. 11.5). На этом рисунке Г. Ф. Михальченко (1989) показала фактически не только динамику возрастания корневого запаса к 45—50 годам, но и его падение с последующим возрастанием. Причем общие закономерности этой тенденции не зависят от климатипов сосны, различаясь только по временной разнице. Характер кривых корневого запаса она объясняла антропогенной, в частности, рекреационной нагрузкой. Но, это далеко не так. Ибо этим насаждениям более 100 лет и запасы стволовой древесины на большинстве участков достигли и даже превышают 400 м3/га.
Волнообразность свойственна отпаду деревьев в лесных культурах. Так, нами на примере культур ели ельника-кисличника с густотой посадки 5 тыс. экз./га на разных фазах роста и развития статикодинамическим методом просматривался общий ход отпада деревьев. До десяти лет отпад наиболее большой в фазе приживания (первые 1—3 года) и объясняется приспособлением растений ели к новой экологической среде. К 10—11 годам отпад почти прекращается, но при наступлении фазы смыкания (смыкания деревьев между рядами) опять увеличивается, достигая максимума в 15 лет. Дальнейшая волна, и как правило наиболее сильная, приурочена к завершению фазы дифференциации, или жердняка (30—45 лет). Отмечено нарастание количества усыхающих стволов в 65—80 лет. За рассматриваемый столетний период наблюдалась четкая волнообразность процесса естественного изреживания в культурах ели, приуроченная к качественно новым перестройкам жизненной структуры древесного сообщества. Подобное явление отмечено и В. В. Галицким (1981), показавшим колебательные изменения биомассы членов растительного сообщества в результате коллективного самоугнетения.
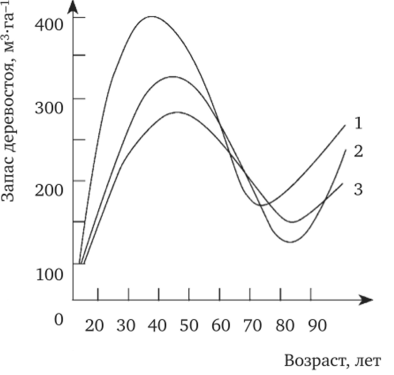
Рис. 29. Кривые изменения корневого запаса сосновых насаждений в условиях Лесной опытной дачи ТСХА.
Климатипы: 1 — местный; 2 — южный; 3 — северный Основная движущая сила отпада — последствия внутривидовой борьбы. До начала очередного интенсивного отпада происходит некоторое торможение ростовых процессов у лесных культур из-за невозможности нормального роста существующего числа стволов. Восстановление нормального роста происходит вслед за отмиранием части стволов, т. е. периоды спада влекут за сбой приливы роста или, вернее, периоды убыстрения роста.
Таким образом, схема концептуальной модели сводится к тому, что весь жизненный цикл искусственного насаждения можно рассматривать как волнообразный, а не плавный процесс роста, то убыстряющийся, то затухающий. Самым первым и вместе с тем самым значительным притуплением роста является «критический возраст» лесокультур в фазе дифференциации (жердняка). В этот момент создается максимальная напряженность в искусственном древостое, вызванная вторжением в жизненную среду каждого растущего культивируемого экземпляра себе подобных индивидуумов. Происходит это по причине ограниченности жизненного пространства растений и ресурсов среды (светового довольствия, содержания элементов питания и влаги в корнеобитаемом слое). Последнее, а также само надземное пространство, занимаемое кронами деревьев, в значительной мере ограничены для каждых условий местопроизрастания и имеют определенную экологическую емкость. Под ней надо понимать способность среды удовлетворять потребность в размещении и питании некоторого количества организмов (Колесниченко, 1976). Это и порождает в определенные жизненные периоды популяции сильную внутривидовую борьбу. Ее следствием является интенсивный отпад части деревьев, т. е. происходит как бы сброс внутренней напряженности искусственного древостоя.
В дальнейшей жизни лесных культур периодически продолжают наступать периоды напряженности и периоды спада напряженности, ибо искусственное насаждение в своем онтогенезе стремится к оптимизации густоты стояния, биологически и эволюционно обусловленной для каждого вида и возрастного этапа. Весь процесс этих волновых приливов с возрастом носит характер затухания. В фазе спелости искусственно созданных древостоев волнообразный характер роста насаждения почти сглажен.
Периодический сброс напряженности и переход в отпад части стволов насаждения просто необходим в силу борьбы за существование части особей. В результате того, что искусственно созданные насаждения являются вместе с тем и самонастраивающимися, саморегулирующимися биологическими системами (по И. И. Шмальгаузену, 1961) и становится возможным то, что периодическая напряженность заменяется отпадом части стволов с последующим усилением роста оставшихся экземпляров.
Торможение роста как насаждения в целом, так и отдельных деревьев при создавшихся неблагоприятных условиях всегда проходит в два этапа: первый — уменьшение прироста по диаметру, второй — по высоте. Поэтому одной из важных характеристик жизненного стояния как дерева, так и древостоя в целом является относительная высота, представляющая собой отношение высоты к диаметру. Этот показатель был введен Я. С. Медведевым в 1884 г. Он считал относительную высоту чувствительным показателем результата светового довольствия (освещенности дерева) и, как следствие, результатом степени напряженности в насаждениях различной сомкнутости. Крайнему угнетению, за которым наступает смерть дерева, соответствует вообще и особенно в молодости (фаза жердняка) весьма высокая относительная высота. Для одного и того же возраста относительные высоты на местах с высшей добротностью всегда ниже, чем на менее добротных.
Относительные высоты с возрастом насаждения уменьшаются. Так как древесные породы имеют неодинаковую потребность в свете и неодинаково способны переносить его недостаток, то и относительные высоты у разных пород при действии одного и того же количества света будут различны. Относительная высота будет иметь наименьшую величину при свободном стоянии дерева и наибольшую в крайне густом стоянии. Например, по В. И. Рубцову (1974) в сосновых культурах относительная высота обычно находится в пределах 90—115, но в случаях с очень высокой густотой стояния, когда деревья испытывают недостаточное количество света, этот показатель существенно повышается.
На основании относительных высот и густот стояния (Мерзленко, 1981) выведен показатель степени напряженности древостоя, который складывается в результате борьбы деревьев за жизненное пространство. Формула имеет следующий вид:
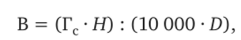
где В — степень напряженности; Гс — густота стояния, шт./га; Н — средняя высота, м; D — средний диаметр, м.
Формула является прежде всего показателем отражения степени внутривидовой борьбы в насаждении.
Учитывая специфику роста и производительности искусственных древостоев, надо все лесохозяйственные мероприятия в деле создания и особенно в деле выращивания лесных культур надо выполнять так, чтобы, выражаясь словами Г. Ф. Морозова (1916), «заставить лесоводов ценить естественные условия роста, обязывая приспособлять свои долженствующие регулировать жизнь сообществ мероприятия к окружающей обстановке».