Регуляция дыхания.
Мышцы, дыхание, выделение, пищеварение, питание

На нейроны дыхательного центра в продолговатом мозге и варолиевом мосту оказывают влияние вышележащие структуры ГМ: гипоталамус, лимбическая система и кора больших полушарий. Гипоталамус участвует в изменениях дыхания, связанных с болевыми раздражениями, физической нагрузкой, терморегуляторными реакциями. Лимбическая система обеспечивает перестройку дыхания при эмоциональном возбуждении, реакциях… Читать ещё >
Регуляция дыхания. Мышцы, дыхание, выделение, пищеварение, питание (реферат, курсовая, диплом, контрольная)
Регуляция дыхания осуществляется структурами СМ и ГМ с помощью сложной системы рефлексов.
Дыхательные движения, обеспечивающие вентиляцию легких, происходят главным образом благодаря сокращениям межреберных мышц и диафрагмы. Эти мышцы иннервируются аксонами мотонейронов, лежащих в соответствующих сегментах СМ. Мотонейроны, в свою очередь, получают команды из дыхательного центра ГМ.
Дыхательный центр не только регулирует ритмические скоординированные сокращения дыхательных мышц, но и приспосабливает дыхание к меняющимся условиям среды и текущим потребностям организма. Большая часть структур дыхательного центра располагается в продолговатом мозге. Его нейроны образуют два отдела: инспираторный (посылает импульсы к мотонейронам, иннервирующим мышцы вдоха) и экспираторный (посылает импульсы к мотонейронам, иннервирующим мышцы выдоха). Оба отдела входят в состав ретикулярной формации продолговатого мозга.
Нейроны инспираторной зоны способны к самопроизвольному периодическому возбуждению (пейсмейкерная активность), и именно они отвечают за периодичность дыхания. При устранении всех афферентных стимулов инспираторные нейроны продолжают в определенном ритме генерировать залпы ПД, которые передаются к дыхательным мышцам. Нарастание импульсации от инспираторных нейронов может быть прервано тормозящими импульсами от пневмотаксического центра и продлено возбуждающими импульсами от апнейстического центра. Эти центры располагаются в структурах варолиева моста: апнейстический центр — в нижних отделах, пневмотаксический — в верхних отделах. Считается, что они участвуют в тонкой настройке дыхания и на базовый ритм не влияют.
Прерывание импульсации от инспираторных нейронов, помимо сигналов от пневмотаксического центра, могут вызывать сигналы от механорецентров, лежащих в стенках легких. Ответом на возбуждение легочных рецепторов растяжения является уменьшение частоты дыхания в результате увеличения времени выдоха. Такая реакция называется рефлексом Геринга — Брейера. Этот рефлекс имеет большое значение для новорожденных, а у взрослого человека срабатывает только при усилении дыхания.
При спокойном дыхании экспираторный отдел дыхательного центра своей активности практически не проявляет. Вентиляция легких обеспечивается активным сокращением инспираторных мышц, которое сменяется пассивным возвратом грудной клетки в исходное состояние. Активный выдох, вызванный активацией экспираторных нейронов, наблюдается исключительно при форсированном дыхании, например, при физической нагрузке.
На нейроны дыхательного центра в продолговатом мозге и варолиевом мосту оказывают влияние вышележащие структуры ГМ: гипоталамус, лимбическая система и кора больших полушарий. Гипоталамус участвует в изменениях дыхания, связанных с болевыми раздражениями, физической нагрузкой, терморегуляторными реакциями. Лимбическая система обеспечивает перестройку дыхания при эмоциональном возбуждении, реакциях страха и ярости. Кора больших полушарий позволяет человеку произвольно изменять частоту и глубину дыхания, а также вызывать его задержку. Помимо этого, кора больших полушарий обеспечивает включение дыхательной системы в поведенческие реакции, речевую функцию и пение. Схема, иллюстрирующая связи дыхательного центра, приведена на рис. 3.15.
Приспособление дыхания к меняющимся условиям среды и потребностям организма обеспечивается сложной системой рецепторов, контролирующих уровень 02, С02, pH, а также состояние легких и воздухоносных путей.
Рецепторы, чувствительные к содержанию в крови 02, С02 и уровню кислотности, сосредоточены в области разветвления общей сонной артерии (каротидный синус) и в дуге аорты. Они входят в состав образований, которые называются, соответственно, каротидными и аортальными тельцами. При снижении содержания 02 в артериальной крови рецепторные клетки этих телец (с помощью медиаторов дофамина и АТФ) активируют окончания чувствительных волокон языкоглоточного и блуждающего нервов. Это приводит к возбуждению инспираторных нейронов и увеличению легочной вентиляции в основном за счет учащения дыхания. При увеличении содержания С02 или снижении pH артериальной крови легочная вентиляция также возрастает, но в основном за счет увеличения глубины дыхания. Рецепторы, контролирующие уровень С02 и pH, помимо крупных артерий располагаются непосредственно в продолговатом мозге на вентральной поверхности (рис. 3.16). При накоплении в ткани мозга углекислого газа или подкислении ликвора эти рецепторы активируются и возбуждают инспираторные нейроны, вызывая усиление дыхания.
Хеморецепторам принадлежит ведущая роль в регуляции дыхания. При этом рецепторы, которые контролируют содержание углекислого газа, более значимы для регуляции, чем рецепторы, которые контролируют уровень кислорода. Это, в первую очередь, связано с лучшей растворимостью С02
в воде и его способностью лучше проходить через биологические мембраны. Примером рефлекторной реакции с участием хеморецепторов являются усиление частоты и возрастание глубины дыхания после его задержки. Поступающий при этом в организм дополнительный объем воздуха точно соответствует объему воздуха, «недополученному» за время задержки (рис. 3.17).
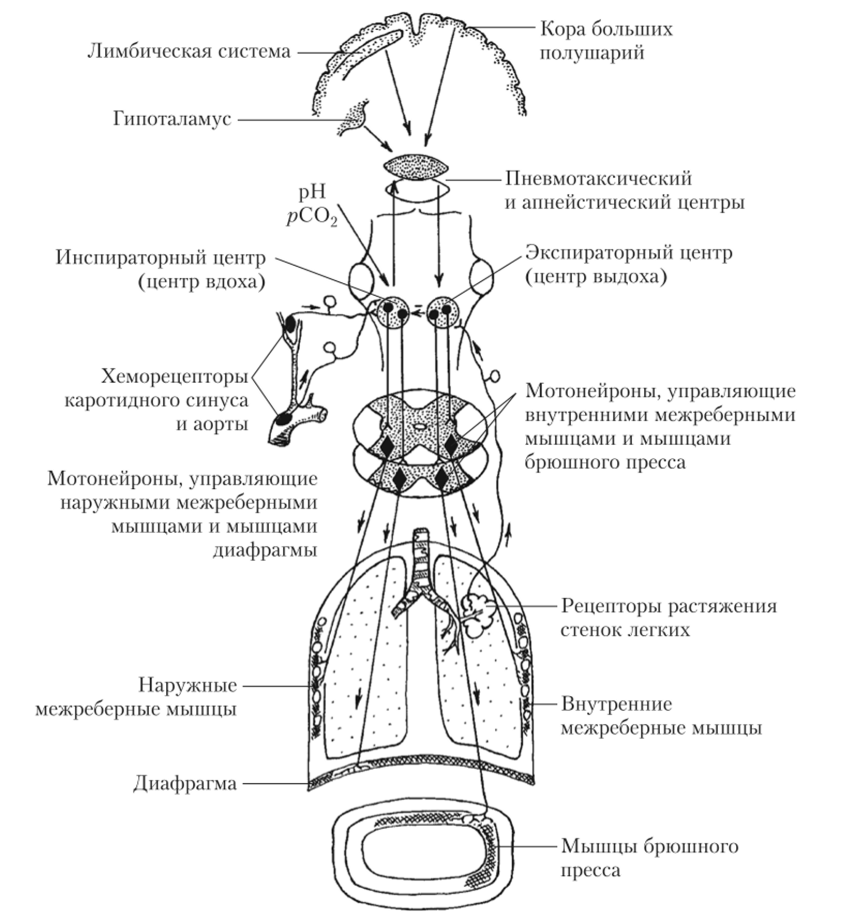
Рис. 3.15. Основные связи дыхательного центра Другим примером дыхательного рефлекса, который «запускается» с хеморецепторов, является уменьшение интенсивности дыхания в ответ на избыточную вентиляцию легких. Объем воздуха, поступившего во время дополнительной вентиляции, равен объему воздуха, который не поступил в организм во время рефлекторного торможения дыхания (см. рис. 3.17).
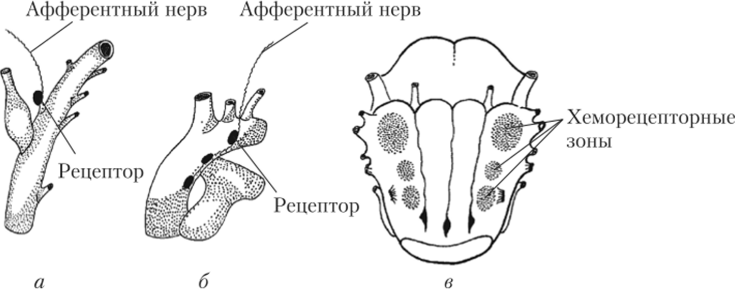
Рис. 3.16. Хеморецепторы, участвующие в регуляции дыхания:
а — каротидные тельца и отходящие афферентные волокна; 6 — аортальные тельца и отходящие афферентные волокна; в — хеморецепторные зоны на вентральной поверхности продолговатого мозга.
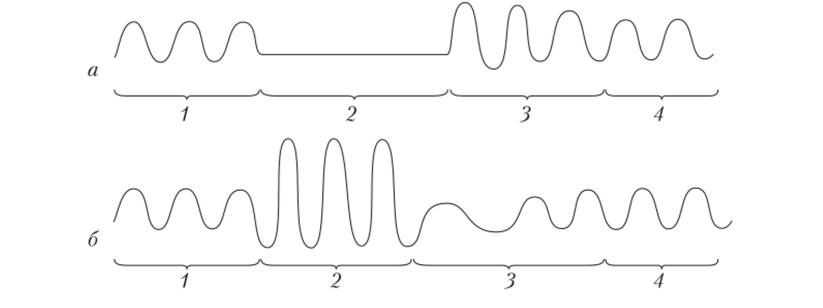
Рис. 3.17. Механограмма дыхательных движений грудной клетки человека при задержке дыхания (а) и гипервентиляции легких (б):
- 1 — исходная норма; 2 — задержка дыхания (а) и гипервентиляция (б)
- 3 — рефлекторная компенсация; 4 — восстановление до исходной нормы
Комплекс хеморецепторов, участвующих в регуляции дыхания, дополняется системой механорецепторов в легких и воздухоносных путях. Эти рецепторы подразделяются на рецепторы растяжения, ирритантные и юкстаальвеолярные.
Рецепторы растяжения располагаются в стенках дыхательных путей и реагируют на растяжение гладких мышц. Это растяжение возрастает при снижении внутриплеврального давления. Благодаря этим рецепторам организм осуществляет контроль над интенсивностью вентиляции легких.
Ирритантные рецепторы обладают свойствами механои хеморецепторов. Они лежат в слизистой оболочке воздухоносных путей и реагируют на изменения интенсивности воздушного потока, действия частиц загрязнителей, скопление слизи или гноя. Эти же рецепторы активируются при действии раздражающих газов (хлора, аммиака, паров фенола и серы) и различных химических веществ и запускают кашель.
Юкстаалъвеолярные рецепторы контролируют объем альвеолярной межклеточной жидкости и кровенаполнение легких. Они расположены в интерстициальном пространстве между стенками альвеол и капилляров. Активация этих рецепторов вызывает учащенное поверхностное дыхание.
Как и хеморецепторы, механорецепторы дыхательных путей запускают многочисленные рефлекторные реакции, рефлекторные дуги которых замыкаются в дыхательном центре продолговатого мозга. Наиболее изученными из этих рефлексов являются следующие:
- 1) инспираторно-тормозящий рефлекс (Геринга — Брейера) — проявляется в торможении вдоха и стимуляции выдоха в ответ на избыточное растяжение легочной стенки;
- 2) кашлевый рефлекс — возникает при раздражении ирритантных рецепторов глотки и нижележащих дыхательных путей. Вначале происходит короткий вдох; затем смыкание голосовых связок и сокращение дыхательных мышц, приводящее к росту подсвязочного давления воздуха. Вслед за этим голосовые связки мгновенно открываются, и воздушная струя с большой скоростью изгоняет из дыхательных путей частицы, вызвавшие раздражение рецепторов (слизь, гной, продукты воспаления или пищевые частицы) в открытый рот;
- 3) рефлекс чихания — развивается подобно кашлевому рефлексу, но вызывается раздражением рецепторов носовых ходов и приводит к изгнанию воздуха через ноздри;
- 4) рефлекс закрытия голосовой щели — проявляется в смыкании голосовых и надгортанных связок в ответ на раздражение рецепторов глотки, гортани и корня языка. Рефлекс защищает дыхательные пути от попадания пищи, жидкости и раздражающих газов;
- 5) рино-бронхиальный рефлекс — запускается раздражением ирритантных рецепторов носовых ходов и проявляется сужением просвета нижних дыхательных путей. У людей с повышенной реактивностью гладких мышц воздухоносных путей пыль и резкие запахи через ирритантные рецепторы могут спровоцировать приступы бронхиальной астмы;
- 6) рефлекс ныряльщика — запускается попаданием жидкости в носовые ходы и останавливает дыхательные движения, блокируя проход жидкости в нижележащие дыхательные пути. Если ныряльщику этот рефлекс может спасти жизнь, то у новорожденных он способен вызвать остановку дыхания. Восстановить нормальный дыхательный ритм у младенца можно, удалив жидкость из носовых ходов.
Таким образом, рассмотренные выше механизмы регуляции дыхания обеспечивают его соответствие потребностям организма и меняющимся условиям среды без участия сознания. Это позволяет «разгрузить» кору больших полушарий и использовать ее возможности для решения более сложных задач.