Модели биологических сообществ

Если в структуре сообщества учитывать движение некоторых биогенных элементов и энергии, то в системе обнаруживаются петли обратной связи. Разлагатели (редуценты) — микробы, грибы, бактерии — в процессе своей жизнедеятельности расщепляют сложные органические соединения (экскременты и мертвую органику) на более простые минеральные вещества, необходимые продуцентам. Образование органической биомассы… Читать ещё >
Модели биологических сообществ (реферат, курсовая, диплом, контрольная)
Структура сообществ
До сих пор мы рассматривали системы уравнений, описывающих взаимодействие двух видов. Однако природные сообщества обладают, как правило, более сложным строением: несколькими уровнями, между которыми существуют разнообразные трофические (пищевые) и топические (не связанные с цепью питания) связи. Структура трофической пирамиды может быть весьма различной в зависимости от климата, почвы, ландшафта, длительности существования ценоза и других факторов.
При анализе биологических сообществ принято строить пищевые или трофические сети, т. е. графы, вершины которых соответствуют видам, входящим в сообщество, а ребра указывают трофические связи между видами. Обычно такие графы — ориентированные: направление дуги между двумя вершинами указывает на тот из видов, который является потребителем другого, т. е. направление дуги совпадает с направлением потока вещества или биомассы в системе.
Часто трофические графы изображают в виде трофических пирамид. В такой пирамиде выделяются трофические уровни — группы видов, между которыми невозможны прямые пищевые связи. Уровней может быть несколько: виды, принадлежащие одному уровню, либо находятся в состоянии конкуренции за жизненные ресурсы, либо совместно используют ресурс. На рис. 4.1 изображен пример двухуровневой трофической пирамиды, взятой из книги Ю. Одума «Основы экологии» (1975). Из 15 видов насекомых (верхний уровень) 5 видов питаются только на одном из двух видов растений, 2 вида — только на втором, в рацион остальных 8 видов насекомых входят оба вида растений. Основные трофические уровни наземных сообществ — это продуценты, или автотрофы, (растения, аккумулирующие энергию света и вещества субстрата) и гетеротрофы: первичные консументы (травоядные) и вторичные консументы (хищники, питающиеся травоядными).
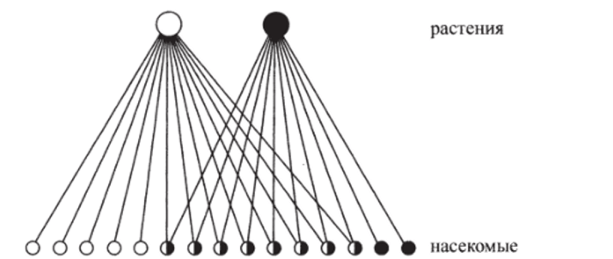
Рис. 4.1. Пример двухуровневой трофической пирамиды (Одум, 1975).
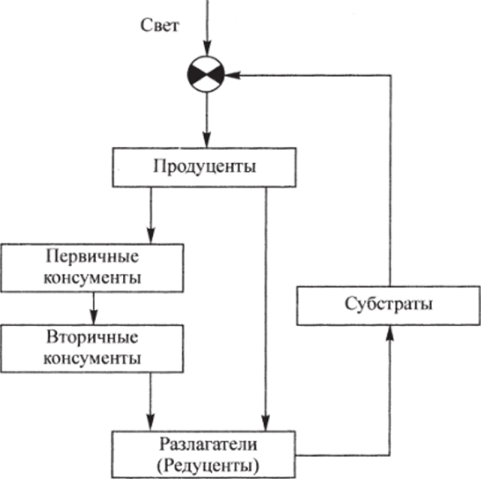
Рис. 4.2. Схема потоков массы и энергии между основными компонентами наземных экосистем.
В некоторых случаях трофическая цепь содержит большее число уровней: например, растения служат пищей насекомым, насекомые поедаются птицами, которые, в свою очередь, служат пищей более крупным хищным птицам.
Если в структуре сообщества учитывать движение некоторых биогенных элементов и энергии, то в системе обнаруживаются петли обратной связи. Разлагатели (редуценты) — микробы, грибы, бактерии — в процессе своей жизнедеятельности расщепляют сложные органические соединения (экскременты и мертвую органику) на более простые минеральные вещества, необходимые продуцентам. Образование органической биомассы происходит в процессе фотосинтеза с использованием солнечного света из углекислого газа и воды, причем необходимы также элементы, поступающие из почвы: азот, фосфор, калий, магний, железо и многие другие микроэлементы.
Общая схема потоков массы и энергии между основными компонентами наземных экосистем изображена на рис. 4.2.
Пусть сообщество состоит из тг видов. Полную структуру парных взаимодействий между видами можно изобразить с помощью матрицы S из п х п элементов. Элемент (г, j) представляет собой знак +, — или 0 и показывает влияние г-ro вида на j-й. Симметричные пары элементов матрицы S указывают на тип парното взаимодействия между видами:
симбиоз или мутуализм. | ||
; | хищник жертва (паразит хозяин). | |
комменсализм. | ||
; | ; | конкуренция. |
—. | аменсализм. | |
нейтрализм. |
Взаимодействие между видами можно характеризовать и при помощи знакового ориентированного графа, который строится, но следующему правилу. Если вид j влияет каким-либо образом на вид г, то проводится ребро г —> j, и ему приписывается знак этого влияния. Например, на рис. 4.3 изображен знаковый граф сообщества, в котором установлены следующие взаимоотношения между видами. Вид 1 — хищник, поедает виды 2 и 3, которые, в свою очередь, питаются видами 4, 5, 6. Виды 4 и 5, а также 5 и 6 связаны отношениями конкуренции. Виды 3, 4. 5 сами ограничивают свою численность, например, по механизму логистического роста (см. раздел 2.1).
В литературе по математической экологии принято считать вольтерровскими моделями сообществ системы вида:
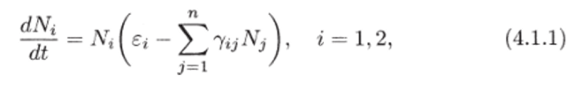
где Si — скорость естественного прироста или смертности г-го вида в отсутствие всех остальных видов, а знак и абсолютная величина 7у (г ф j) отражают соответственно характер и интенсивность влияния j-ro вида на г-й вид, показатель внутривидового взаимодействия для i-ro вида. Матрицу Г = ||7о11? отражающую структуру связей сообщества, называют матрицей сообщества.
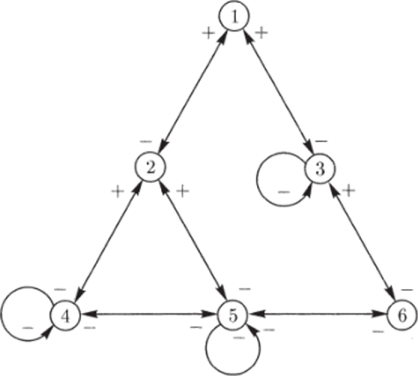
Рис. 4.3. Пример знакоориентированного сообщества (Свирежев, Логофет, 1978).
С введенной выше знаковой матрицей S она связана соотношением.
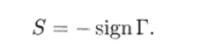
Гипотеза Водьтерра, на основе которой построена система (4.1.1), предполагает локальный принцип описания взаимодействия видов — принцип «встреч», ведущий свое происхождение из статистической физики. Вольтерровские уравнения могут быть также получены из чисто экологических предпосылок (Свирежев, 1976).
Рассмотрим сообщество, структура которого изображена на рис. 4.3. Компоненты сообщества разобьем на три основные группы.
Продуценты с биомассами (или концентрациями) х (г = = 1,2, …, га) это в основном зеленые растения.
Консументы с концентрациями yj (j = 1,2, …, п). К этой группе отнесем животных, пожирающих другие организмы, и разлагателей, расщепляющих мертвую органику на простые вещества, которые используются продуцентами.
Субстраты с концентрациями с* (к = 1,2, …, р). Это абиотические вещества (в основном продукты жизнедеятельности консументов), используемые продуцентами.
Составим уравнения, отражающие баланс масс каждого из этих компонентов:
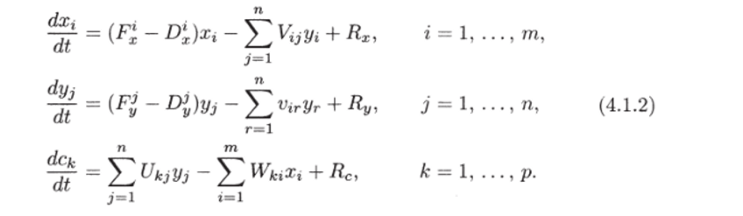
Здесь F1, D1 — функции рождаемости и смертности продуцентов и консументов; Vij — функция выедания, описывающая скорость потребления биомассы г-го вида-продуцента единицей биомассы j-го вида-консу мента; Vjr — функция выедания j-го вида г-м (среди консументов); Ukj — интенсивность производства к-го субстрата j-м консу ментом; Wki — интенсивность потребления к-го субстрата г-м продуцентом; Rx = Ry = Rc — сумма входных и выходных потоков соответствующих компонент. В общем случае все эти функции зависят от параметров внешней среды.
Конкретизируем функции, входящие в обобщенную модель (4.1.2). Пусть рождаемость г-ro вида — продукта FJ — зависит только от интенсивности светового потока I и количества потребляемых субстратов в форме 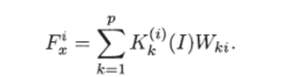
Здесь — стехиометрические коэффициенты, показывающие долю к-го субстрата, идущего на построение биомассы ?-го продуцента.
Для скорости прироста ji-ro консумента аналогично получаем FjJ =.
= + 12 Pr^Vrj, где К^ Prj) — стехиометрические коэффи;
1=1 г=1.
циенты взаимодействия консументов с продуцентами и хищничества среди консументов.
Сделаем дальнейшие упрощающие предположения. Пусть рождаемость продуцентов не лимитирована ни светом, ни минеральным питанием, а ограничена лишь чисто физиологическими пределами. Тогда Fx = const. Субстраты в этом случае не оказывают влияния на динамику остальных компонент, поэтому в модели можно рассматривать только виды-продуценты и виды-консументы и третье уравнение из системы (4.1.2) исключить. Предположим также, что система замкнутая и Rx = Ry = Rc = 0.
Пусть функция смертности линейно зависит от численности соответствующих видов:
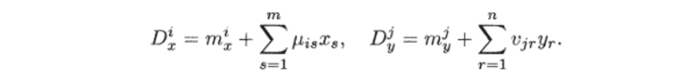
Здесь тгх, тj — коэффициенты естественной смертности продуцентов и консументов, /i#, Vjj описывают интенсивность внутривидовой конкуренции.
Пусть функция выедания также линейно зависит от концентрации (биомассы) выедаемого вида:
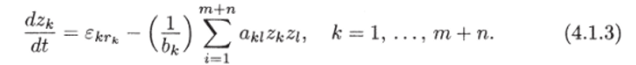
Здесь.
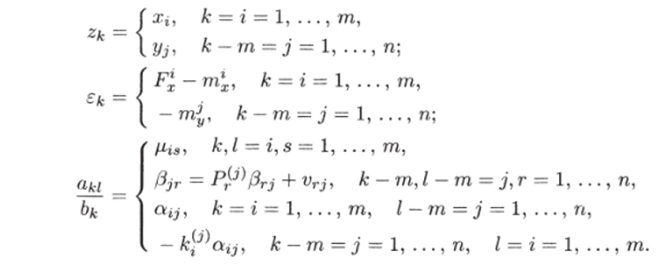
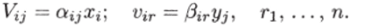
Учитывая сделанные предположения, модель (4.1.2) можно записать в виде Система (4.1.3) путем преобразования переменных TV* = bkZk и введением матрицы с элементами 7*./ = (iki/bkbi может быть записана в виде (4.1.1) и представляет собой самый общий вид вольтерровских моделей.
Таким образом, рассмотрение балансовых соотношений массы в бжл’еоценозе приводит к уравнениям, сходным по форме с уравнениями Вольтерра. Поэтому важные выводы, сделанные Вольтерра из математического анализа своих систем, правильны и актуатьны с точки зрения современной экологической науки.
При моделировании каждой конкретной экосистемы требуется внимательное изучение характера взаимодействий видов, как находящихся на одном трофическом уровне, так и вступающих в пищевые взаимодействия. При этом, как и в случае взаимодействия двух видов, функции рождаемости, смертности и выедания могут носить более сложный характер, чем это предусматривается гипотезами Вольтерра. Часто система уравнений, описывающих динамику видов, так сложна, что решение и исследование устойчивости стационарных состояний такой системы может быть выполнено только с помощью компьютерных программ. Однако некоторые общие выводы можно сделать при исследовании упрощенных систем.