Организация жизни в биосфере

Биосферы С другой стороны, определенная популяция, вступая в трофические (греч. trophe — пища, питание) и иные отношения с популяциями других видов, является неотъемлемой составной частью экосистемы и биосферы в целом, в сложной системе передачи и преобразования вещества и энергии популяция выступает как функциональный элемент. Такой подход отражает положение популяции как составной части… Читать ещё >
Организация жизни в биосфере (реферат, курсовая, диплом, контрольная)
Популяции
Закон толерантности Шелфорда рассматривает действие экологического фактора на единичный организм. При изучении воздействия какоголибо фактора па группу организмов одного вида оказывается, что для каждого организма оптимальная область, величина толерантности и область угнетения (см. рис. 3.8) несколько различаются.
На рис. 4.1 приведена упомянутая зависимость для трех организмов одного вида. Рассмотрим воздействие экологического фактора определенной величины K,)(j}1 на три организма одного вида, для первого организма К^ф! будет соответствовать оптимальной области, для второго и третьего — области угнетения. Если величина экологического фактора возрастет до Кэф2, то третий организм погибнет, а выживут первый и второй.
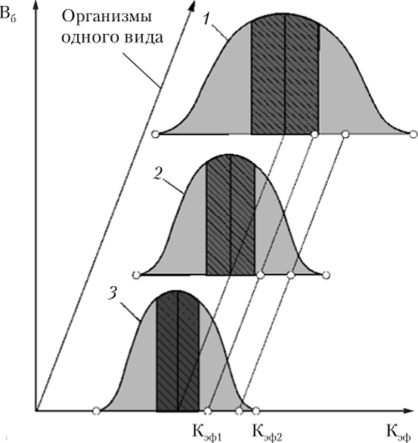
Рис. 4.1. Воздействие экологического фактора на группу организмов.
(организмы: 1, 2, 3)
Таким образом, все живые организмы, для того чтобы обеспечить устойчивое существование и воспроизводство вида в условиях изменяющихся экологических факторов, должны существовать группами, которые называются популяциями (лат. populus — народ, население). В научной литературе существует немало понятий термина «популяция». Микробиологи, генетики, биологи вкладывают свои, зачастую различные представления в этот термин. Такие подходы связаны с двойственностью положения популяции в иерархии биологических систем, отражающих различные уровни и структуры организации живого вещества. С одной стороны, популяция является составной частью гепетико-эволюционного ряда и выступает как структурная единица вида, ее основная функция заключается в обеспечении устойчивого эволюционного развития вида (рис. 4.2).

Рис. 4.2. Положение популяции в структуре биологических систем.
биосферы С другой стороны, определенная популяция, вступая в трофические (греч. trophe — пища, питание) и иные отношения с популяциями других видов, является неотъемлемой составной частью экосистемы и биосферы в целом, в сложной системе передачи и преобразования вещества и энергии популяция выступает как функциональный элемент. Такой подход отражает положение популяции как составной части функционально-энергетического ряда. Выполнение этой функции основывается на свойствах адаптивности популяции, способности поддерживать устойчивое динамическое равновесие в условиях изменяющихся экологических факторов. В результате обеспечивается стабильное участие вида в биогеохимических циклах.
С учетом изложенного наиболее полной, на наш взгляд, является следующая формулировка. Популяция — совокупность особей одного биологического вида, населяющих пространство с относительно однородными экологическими условиями, имеющих общий генофонд и возможность свободно скрещиваться.
Популяция (homo) — население, заселяющее определенную территорию в течение пяти-шести поколений с определенным климатогеографическим, микроэлементами, радиационным составом окружающей среды.
Любая популяция занимает определенное место в пространстве таким образом, чтобы это позволяло наиболее эффективно использовать ресурсы окружающей среды и обеспечивало устойчивые внутрипопуляционные взаимоотношения особей и их групп.
Пространственное размещение особей в популяции может носить равномерный, случайный и групповой характер (рис. 4.3). При равномерном размещении особи распределены в пространстве примерно на одинаковых расстояниях друг от друга. Такое размещение чаще всего встречается в популяциях растений (хвойные деревья в тайге). Этот тип размещения позволяет наиболее полно использовать ресурсы окружающей среды.
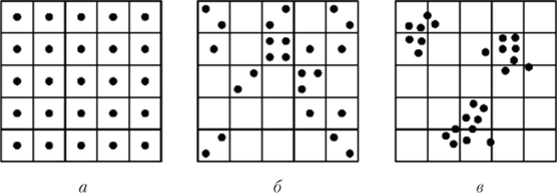
Рис. 43. Типы пространственного распределения особей в популяции:
а — равномерное; б — случайное; в — групповое Случайное размещение встречается в природе наиболее часто и наблюдается как среди растений, так и среди животных. Особи при этом распределены неравномерно, расстояния между ними неодинаковы, поэтому их встречи носят достаточно случайный характер.
Групповое размещение выражается в образовании группировок особей, между которыми имеются значительные незаселенные территории (многие млекопитающие, колонии насекомых и птиц, осоковые кочки на болотах).
Как правило, особи одной и той же популяции, находящиеся на большом географическом пространстве, образуют смешанное из отмеченных трех типов пространственное распределение.
При рассмотрении пространственной структуры популяций животных важное значение имеет степень привязанности к территории. При этом выделяют два основных образа жизни: оседлый и кочевой.
При оседлом образе жизни, как правило, наблюдается интенсивный тип использования территории. Отдельные особи или группировки в этом случае в течение длительного времени эксплуатируют ресурсы окружающей среды на достаточно ограниченном пространстве (группы бобров, рыжих полевок, рептилий).
Для популяций, ведущих кочевой образ жизни, характерен экстенсивный тип использования территории. При этом кормовые ресурсы и питьевая вода используются обычно группами особей, постоянно перемещающихся в пределах обширных территорий (стада северных оленей или африканских слонов).
Поддержание пространственной структуры у популяций животных обеспечивается за счет агрессии и маркирования территории. Преимущество обитания на постоянной территории реализуется путем использования его только отдельной особью или группой особей. Наиболее выраженной формой территориальных внутривидовых взаимоотношений является эволюционно сформировавшийся стереотип территориальной агрессии. Агрессивное поведение направлено на удаление особей своего вида, проникших на чужую территорию. Исследованиями установлено, что при таких конфликтах победителем чаще всего выходит особь — «хозяин» участка. Такой стереотип поведения характерен для популяций, ведущих оседлый образ жизни, причем во многих случаях территориальная агрессия носит демонстративный характер без физического контакта между соперниками.
Маркировка территории не связана с агрессивным способом закрепления участка за особью. Способы маркировки разнообразны: визуальный, акустический, запаховый. Так, у коралловых рыб яркая окраска является достаточным сигналом занятости территории. Самцы тюленя маркируют подводный участок вокруг своего убежища с помощью издаваемых трелей. Акустическая маркировка ярко выражена у птиц. В сложной мелодии песни имеется фрагмент, означающий территориальный сигнал. У большинства млекопитающих маркировка территории осуществляется с помощью запаховых меток. Для этих целей используется моча, экскременты или секреты особых желез. Эффективность такого способа поддержания пространственной структуры популяции связана с наличием врожденной негативной реакции на чужой территориальный сигнал.
Пространственное структурирование популяции позволяет использовать максимальное количество ресурсов среды путем снижения конкуренции. Вместе с тем пространственная дифференциация ограничивается необходимостью интеграции особей в пространстве, так как без интеграции невозможна реализация важнейших общепопуляционных функций. К ним относятся воспроизводство, регуляция численности, общепопуляционные реакции на внешние з’словия, взаимоотношения с популяциями других видов, т. е. популяция также должна быть структурирована и функциональна.
Основу функционального структурирования популяции составляют два взаимосвязанных процесса: непрерывный обмен информацией о месте нахождения отдельных особей и группировок и эволюционно сформированный стереотип поведения, стимулирующий особь к поиску и поддержанию контактов с себе подобными.
Информация о присутствии особей своего вида достаточно разнообразна. Это звуковые и ультразвуковые сигналы, визуальное восприятие, запахи. У некоторых рыб источниками информации являются электрические разряды, у ряда видов беспозвоночных — механические колебания воды. Весь объем информации в виде различных сигналов, изменений растительности и рельефа и других следов жизнедеятельности образуют биологическое сигнальное поле, которое является информационной основой интеграционных процессов в популяции. Так, ящерицы в лабораторных условиях активно стремятся присоединиться к особям своего вида, а при их отсутствии — к особям близкого вида [61, с. 266].
В основе поддержания функциональной структуры популяции лежит система соподчинения животных, при этом прослеживается довольно сложная иерархическая структура взаимоотношений. У птиц в определении ранга зачастую решающее значение имеет окраска, часто связанная с возрастом. У стадных животных важную роль в становлении вожака играют генетически определенные особенности физиологии и высшей нервной деятельности.
Каждая популяция характеризуется различной степенью генетического разнообразия особей. Совокупность генов, которыми обладают особи популяции, называется генофондом популяции. Чем более разнороден и богат генофонд популяции, тем выше ее экологическая пластичность. Особи, входящие в состав популяции, неравноценны по своим функциям, генетическому вкладу и соответственно по своим индивидуальным свойствам. В этом заключается одна из особенностей функционального структурирования популяции.
Совокупность всех генов, находящихся в хромосомах организма, называется генотипом. Это зашифрованная программа, управляющая процессом развития и обеспечения жизнедеятельности организма. На развитие и жизнедеятельность организма оказывают воздействие экологические факторы, поэтому в любой популяции отсутствуют особи, абсолютно идентичные по своим признакам и свойствам. Совокупность всех признаков организма, представляющих собой результат взаимодействия генотипа с окружающей средой, называют фенотипом.
Универсальным свойством всего живого, от микроорганизмов до высших растений и животных, является способность к мутациям. Мутации — это наследуемые изменения генетических программ, приводящие к изменению определенных признаков организма. Они проявляются в фенотипе взрослого организма, поскольку индивидуальная генетическая программа его развития от момента оплодотворения и до смерти отличается от генетических программ других особей популяции.
Важнейшим фактором эволюции популяций является индивидуальный естественный отбор — отбор наиболее приспособленных к среде особей, т.е. тех особей, у которых мутации повысили устойчивость организма к экологическим факторам среды.
Изменения условий окружающей среды вызывают соответствующие адаптивные перемены и в пространственно-функциональной структуре популяций. Так, пятнистые гиены образуют кланы до 100 особей в том случае, когда передвижение объектов их охоты (копытных животных), например в предгорьях, ограничено. На равнинах, где гиены должны перемещаться за стадами копытных животных на большие расстояния, таких кланов не отмечено.
Таким образом, пространственное и функциональное структурирование популяции обеспечивает ее бесперебойное функционирование.
Естественные популяции не являются статически неизменными системами. в любой популяции происходят сложные динамические процессы. Наиболее важными показателями, характеризующими эти процессы, являются рождаемость и смертность.
Рождаемость — это интенсивность восполнения популяции за счет размножения. В экологии различают максимальную рождаемость — рождаемость при оптимальных значениях экологических факторов, воздействующих на организм. В этом случае размножение организмов сдерживается их физиологическими особенностями. Как уже отмечалось в параграфе 2.4, теоретическая скорость размножения различных видов может быть достаточно высокой. Для определенной популяции величина максимальной рождаемости колеблется в незначительных пределах, т. е. является практически постоянной.
В реальных условиях реализованная (экологическая) рождаемость
значительно ниже, поскольку вся совокупность экологических факторов, воздействующих на организм, никогда нс может быть оптимальной. Для оценки реализованной рождаемости используют индекс рождаемости Rh, который равен отношению числа новорожденных особей п за единицу времени к количеству особей в популяции N.
Смертность — это интенсивность гибели особей в популяции. Характеризуется индексом смертности Rd, равным отношению числа умерших или погибших особей т за единицу времени к количеству особей в популяции N. Смертность в популяции может быть физиологической, когда особи реализуют весь генетически определенный цикл жизнедеятельности организма, и экологической, когда часть особей умирает под воздействием факторов окружающей среды, не достигнув предельного возраста.
Вместе с рождаемостью смертность определяет характер изменения численности популяции. При Rb > Rd наблюдается рост популяции, при Rh < Rd — происходит снижение численности особей в популяции. Если Rd = Rb то происходит стабилизация численности.
Экологически реальный рост популяции выражается логистическим уравнением Ферхульста — Пирла:

где г — скорость роста популяции, обусловленная физиологическими особенностями организма; t — время; N — количество особей в популяции; К — емкость среды (максимально возможная численность популяции в данной экосистеме).
Множитель (К — N)/K характеризует сопротивление окружающей среды росту популяции (рис. 4.4). При отсутствии сопротивления среды полностью реализуются физиологические возможности организма в области размножения, т. е. реализуется биологический потенциал популяции, который описывается уравнением экспоненциального роста dN/dt = rN0 (N0 — исходная численность популяции в начальный момент времени).

Рис. 4.4. Рост численности популяции:
1 — экспоненциальная кривая; 2 — логистическая кривая Экспоненциальный рост популяции в биосфере Земли, как правило, не наблюдается, поскольку имеется ряд лимитирующих экологических факторов. Только рост популяции людей в XX—XXI вв. пока близок к экспоненте, так как благодаря достижениям науки и техники человечеству удалось существенно улучшить свою экологическую нишу и приблизить значения экологических факторов к оптимальным.
Экспериментальная проверка уравнения Ферхульста — Пирла выполнена советским биологом Г. Ф. Гаузе в 1934 г. Он исследовал размножение реснитчатых инфузорий в лабораторных условиях, при этом кривая роста численности инфузорий практически соответствовала логистической кривой.
Наблюдения показывают, что численность большинства видов может быть постоянной достаточно длительное время. В эволюционных масштабах такое равновесие сформировалось из-за согласованности рождаемости и смертности, называемой регуляцией численности популяции. Так, кладка рыбы-луны содержит до 300 млн икринок, но эта икра доступна для поедания другими животными, в то же время акулы откладывают всего несколько яиц в сезон, но эти яйца защищены плотной оболочкой. Похожие явления наблюдаются и среди земноводных: зеленая жаба, не заботящаяся о потомстве, откладывает за сезон до 12 тыс. икринок, тогда как жаба-повитуха, наматывающая свою икру на задние лапы, — всего около 150 икринок.
Многочисленными исследованиями установлено: чем выше забота о потомстве, тем ниже у таких видов смертность в раннем возрасте и соответственно ниже рождаемость. Кроме того, у видов, выкармливающих свое потомство, рождаемость в большей степени зависит от возможности обеспечить выводок кормом. Также известно, что рождаемость связана со средней продолжительностью жизни, свойственной данному виду: у долгоживущих видов рождаемость ниже [61, с. 312—325].
В современной экологии выделяют две группы факторов, обеспечивающих регуляцию численности популяции: 1)факторы, не зависящие от численности популяции; 2) факторы, связанные с численностью популяции.
К первой группе относится комплекс абиотических экологических факторов, которые в своем воздействии на организмы реализуются через составляющие климата и погоды. Эти факторы действуют на уровне организма, поэтому результат их воздействия не зависит от таких показателей популяции, как численность и плотность населения. Организмы могут только адаптироваться к этим факторам, но не в состоянии их изменить. Так, если наблюдается суровая зима при небольшой толщине снежного покрова, то к весне численность в популяциях мелких грызунов окажется достаточно низкой, даже если к началу зимы плотность была высокой. Аналогичная закономерность характерна и для растений.
При некоторых условиях абиотические факторы могут действовать и косвенно, например через изменение кормовых и других условий. Так, при длительной засухе наблюдается пересыхание водоемов, которое приводит как к массовой гибели водных организмов, так и к уменьшению растительных кормовых ресурсов.
Климатические факторы, носящие циклический характер, также могут быть причиной изменений численности. Установлено совпадение циклов изменения численности грызунов с 11-летними циклами солнечной активности.
Характер влияния факторов, зависящих от численности популяции (факторы «авторегуляции»), существенно отличается от уже рассмотренных. Это выражается во взаимовлиянии популяций различных видов друг на друга. Регуляция при этом осуществляется по кибернетическому принципу регулирования системы с обратной связью. Экспериментальные исследования в этой области показывают, что популяцию при этом можно рассматривать как колебательное звено с отрицательной обратной связью, поэтому численность популяции колеблется возле оптимального значения в данных условиях.
При формировании циклов численности важное значение имеют отношения потребителя к пище. Высокая обеспеченность пищей ведет к росту рождаемости и снижению смертности, в результате численность популяции растет. Это в свою очередь приводит к увеличению потребления кормовых ресурсов и снижению их объемов. Так, выедание растительности ведет не только к сокращению ее количества, но и к изменению ее качества — снижению удельного содержания фосфора, кальция, белка и т. д. При этом также снижаются защитные функции растительности как места укрытия от хищников. В результате с некоторым запаздыванием (как это и должно быть в автоматической системе регулирования с колебательным звеном) численность популяции уменьшается, снижается давление на растительность и ее объем возрастает. Численность популяции начинает возрастать, и цикл повторяется снова. Классическим примером таких взаимоотношений являются циклы изменений численности норвежского лемминга, отличающиеся катастрофическими выеданиями тундровой растительности. В результате происходит массовая миграция и гибель большого числа этих животных.
Наиболее четко циклы изменения численности прослеживаются в условиях взаимоотношений хищников с жертвами. При этом они оказывают взаимное влияние на численность и плотность особей в обеих популяциях. При отсутствии влияния других факторов происходят повторяющиеся колебания численности обоих популяций, причем изменения численности хищника отстают по фазе от колебаний численности жертвы (рис. 4.5).
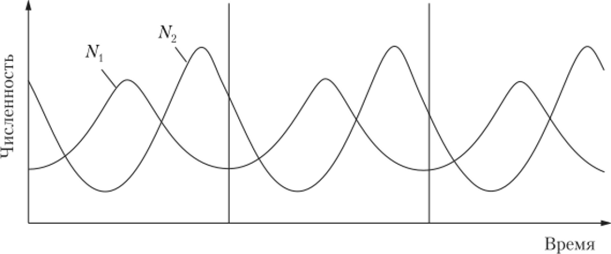
Рис. 4.5. Взаимосвязанные колебания численности популяций жертвы (JVt) и хищника (N2) по модели Лотки — Вольтерра.
Интересны эксперименты, поставленные замечательным биологом Г. Ф. Гаузе. В лабораторных условиях, исключающих влияние других факторов, исследовано взаимодействие хищных инфузорий (хищник) с парамециями (жертва). В однородной среде хищники съедали всех своих жертв и вымирали сами, т. е. реализовывался всего один цикл. При наличии укрытий для жертвы хищники вымирали, а рост численности жертв практически совпадал с логистической кривой. Последовательные колебания численности обеих популяций наблюдались при регулярном добавлении к ним особей обоих видов, что имитировало «иммиграцию» новых особей в области пониженной плотности. Многие экологи считают, что эксперименты Г. Ф. Гаузе не подтвердили теоретическую модель Лотки — Вольтерра. Авторы не могут с этим согласиться, так как, во-первых, был экспериментально реализован полный цикл, в котором достаточно четко прослеживались гармонические колебания численности обеих популяций с отставанием по фазе колебания численности хищника. Это подтверждает теоретическую предпосылку, что взаимодействия в системе «хищник — жертва» носят колебательный характер с отставанием по фазе одного из процессов.
Во-вторых, в реальных условиях при уменьшении численности жертвы хищники расширяют круг поисков или переключаются на жертвы из других популяций, уменьшая тем самым давление на первую популяцию жертвы, давая ей возможность увеличить численность.
Кроме того, любая имитационная модель в любой науке всегда отличается в худшую сторону от реального объекта. На ней можно подтвердить только основные положения, относящиеся к реальным процессам, поэтому, на наш взгляд, эксперименты Г. Ф. Гаузе в достаточной степени подтверждают выводы Лотки — Вольтерра.
Динамика численности в системах типа «паразит — хозяин» примерно соответствует колебаниям численности в рассмотренной системе.
На численность животных существенное влияние оказывают эпизоотии (массовые инфекционные заболевания животных). При увеличении плотности особей в популяции облегчается передача возбудителей болезней от одной особи к другой и повышается смертность. Снижение численности особей до определенного значения приводит к прекращению переноса возбудителей болезней, при этом в выжившей части популяции сохраняются особи, приобретшие иммунитет. Они являются хранителями возбудителя до следующей вспышки.
Схожие явления наблюдаются и в растительном мире при эпифиотии (массовые инфекционные заболевания растений).
Каждая популяция имеет определенное соотношение особей различного возраста. В популяции, как правило, можно выделить три экологические группы: пререпродуктивную (довоспроизводящую), репродуктивную (воспроизводящую) и пострепродуктивную (послевоспроизводяищю).
Практически для любой популяции темпы ее роста в каждый момент времени определяются долей особей, находящихся в репродуктивном возрасте. Количество нреренродуктивных особей отражает потенциальные возможности по изменению своей численности в ближайшем будущем.
Возрастные спектры для различных популяций неодинаковы (рис. 4.6). Для видов, смертность которых незначительно зависит от внешних факторов, график выживания вначале имеет вид пологой кривой (рис. 4.6, линия 1), а при достижении возраста физиологической смерти наблюдается резкое ее снижение. В биосфере таких видов немного — в основном насекомые с непродолжительным сроком жизни. Так, у поденок личиночное развитие (пререпродуктивная стадия) достигает нескольких лет, репродуктивный период — несколько дней, а пострепродуктивный — практически отсутствует [61, с. 330—333].
При равномерном распределении смертности по возрастам график выживания приближается к диагонально снижающейся прямой линии (рис. 4.6, линия 2), что характерно для высокоразвитых млекопитающих.
Для многих видов повышенная смертность наблюдается в пререпродуктивный период, соответственно кривая смертности при этом резко падает (рис. 4.6, линия 3). Затем наблюдается плавное снижение кривой, характеризующее равномерную смертность особей в оставшихся периодах.
В результате хозяйственной деятельности человека образовались природно-антропогенные популяции. Так, популяция колорадского жука связана с выращиванием картофеля. В местах обитания человека, размещения отходов жизнедеятельности возникают популяции грызунов и насекомых (мухи, тараканы, мыши, крысы и т. д.). Они могут быть возбудителями и переносчиками опасных инфекционных заболеваний, т. е. выступать в качестве опасных экологических факторов.

Рис. 4.6. Возрастные спектры различных популяций.
В последние 100—150 лет человек оказывает существенное влияние на многие природные популяции, причем, как правило, в сторону уменьшения их численности. При низкой численности популяций уменьшается вероятность скрещивания, ухудшаются показатели естественного отбора. Это относится как к животному, так и к растительному миру.
Виды, находящиеся под угрозой исчезновения, заносят в особые перечни — Красные книги. Эти виды находятся под особой охраной государств и международных организаций. Начало созданию Красных книг положено в середине прошлого века. В 1948 г. по инициативе ЮНЕСКО был образован Международный союз охраны природы и природных ресурсов, который провел большую работу по выяснению численности тех видов животных и растений, которым угрожает исчезновение. К настоящему времени издано пять томов международной Красной книги.
В СССР Красная книга была учреждена в 1974 г., первое ее издание состоялось в 1978 г., второе — в 1985 г.
Красная книга Российской Федерации вышла в свет в 2001 г. Она представляет собой 860 страниц текста, иллюстрирована цветными изображениями всех занесенных в нее животных и картами их ареалов. Всего в Красную книгу Российской Федерации занесено 415 видов и подвидов, в том числе 155 таксонов беспозвоночных и 260 — позвоночных животных. Общий список по сравнению с Красной книгой РСФСР увеличен на 73%, причем главным образом за счет видов и подвидов беспозвоночных животных (объем группы увеличен в 3 раза), а также рыб и рыбообразных (в 4 раза).
В 1979 г. постановлением Совета Министров БССР учреждена Красная книга Белорусской ССР. В первое издание данной книги включено 80 видов животных и 85 видов сосудистых растений. Второе издание Красной книги Республики Беларусь состоялось в 1993 г.: охране подлежали 182 вида животных, 180 видов растений, 17 видов грибов и 17 видов лишайников, третье издание Красной книги Беларуси состоялось в 2005 г.