Дифференциальная активность генов

Широко распространен механизм регуляции активности ферментов, основанный на присоединении к ним молекул-эффекторов. Чаще всего в роли эффекторов выступают конечные продукты цепей биосинтеза, которые связываются с первым или с одним из первых ферментов данного метаболического пути и подавляют его активность, тем самым выключая всю цепь синтеза. Это ингибирование конечным продуктом, благодаря… Читать ещё >
Дифференциальная активность генов (реферат, курсовая, диплом, контрольная)
Дифференциальная экспрессия генов, т. е. регуляция их активности в зависимости от сигналов, поступающих извне, может происходить на уровне любого известного матричного процесса: репликации, транскрипции, трансляции, а также в процессе созревания иРНК и полипептидных цепей, образующихся в результате трансляции.
Дифференциальная репликация отдельных участков генетического материала известна у прокариот и эукариот. Индукция и последующая репликация профага X представляют собой пример дифференциальной репликации у бактерий. Амплификацию экстрахромосомных копий ДНК, кодирующей рРНК, наблюдали в ядрышках ооцитов многих животных, а также при мегаспорогенезе растений. Амплификация рДНК заключается в том, что одна из ее копий, содержащая многократные повторы генов, кодирующих рРНК, покидает хромосому — область ядрышкового организатора, затем многократно реплицируется по механизму катящегося кольца. Этим достигается усиленный синтез рибосом в ооците, обеспечивающий ранние этапы развития после оплодотворения. Известно, что сперматозоид вносит в зиготу только ядерный материал и первые стадии дробления вплоть до гаструлы обеспечиваются цитоплазмой, а следовательно, и рибосомами яйцеклетки.
В гигантских хромосомах слюнных желез двукрылых наблюдается большая политенизация отдельных участков хромосом. Само образование политенных хромосом указывает на то, что репликация в различных соматических клетках происходит неодинаково. Об этом же свидетельствует и сравнение ретиконов — единиц репликации различных соматических клеток. Размеры репликонов в ходе дифференцировки тканей изменяются.
Дифференциальная транскрипция генов в онтогенезе хорошо заметна при образовании хромосом типа ламповых щеток, петли которых, возникающие на стадии диплотены, активно транскрибируются, что хорошо видно на электронно-микроскопических препаратах ооцитов амфибий и птиц.
Другой яркий пример дифференциальной транскрипции связан с образованием так называемых пуфов, или колец Бальбиани, в гигантских хромосомах двукрылых. Пуфы — это характерные вздутия определенных дисков политенных хромосом, образующиеся в результате локальной декомпактизации в них ДНК, сопровождающейся активной транскрипцией, на что указывает интенсивное включение 3Н-уридина в районе пуфов на препаратах политенных хромосом (рис. 10.3).
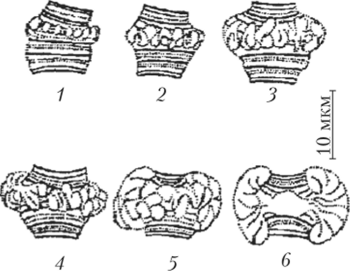
Рис. 10.3. Образование пуфа, или кольца Бальбиани, в политенной хромосоме на последовательных стадиях развития (1−6) личинки хирономуса Пуфирование тех или иных дисков характерно для стадии развития личинки. Образование и исчезновение пуфов регулирует внутренняя среда организма в соответствии со стадией развития. Если слюнные железы личинки дрозофилы пересаживать более молодым или более старым личинкам, то картина распределения пуфов меняется в соответствии с той, которая характерна для возраста реципиента (рис. 10.4).
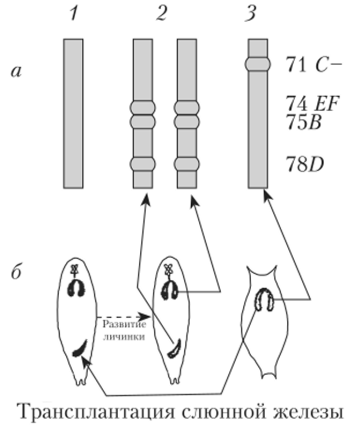
Рис. 11.4. Изменение характера пуфирования в хромосоме D. melanogaster в результате пересадок слюнных желез более молодым личинкам: а — гтуфы в 3-й хромосоме личинок разных возрастов (1−3). Справа — номера дисков на цитологической карте; б — трансплантация слюнной железы из более старой личинки в молодую и сравнение характера пуфирования «своих» и «чужих» хромосом при последующем развитии Одним из важных регуляторов образования пуфов и, следовательно, дифференциальной транскрипции генов у насекомых являются стероидные гормоны, в частности гормон линьки — экдизон.
Изменение структуры хроматина, его декомпактизация, наблюдаемая при образовании пуфов, также является одним из условий, обеспечивающих дифференциальную активность генов.
Дифференциальная трансляция, т. е. синтез белка только на определенных иРНК или регуляция синтеза белка на одной и той же иРНК, показана для РНК-содержащих бактериофагов Е. coli, а также при синтезе глобинов на стабильных иРНК безъядерных ретикулоцитов млекопитающих. Избыток гемина стимулирует синтез глобина. Гемин инактивирует белок, который репрессирует, т. е. «запрещает» синтез аи р-цепей глобина. Некоторые фракции тРНК играют роль модуляторов, задающих темп трансляции; тРНК-модуляторы служат лимитирующим фактором в трансляции, «узнавая» какой-либо уникальный кодон иРНК. Гипотеза модулятора была предложена в начале 1960;х гг. Г. Стентом.
Возможность дифференциальной трансляции основывается на существовании стабильных иРНК, а также на сохранении иРНК в цитоплазме в виде информосом — комплекса иРНК с белками, открытого в 1966 г. А. С. Спириным и др.
Дифференциальное созревание продуктов транскрипции и трансляции. Созревание транскриптов подразумевает модификацию их отдельных оснований и сплайсинг про-иРНК. Несколько вариантов сплайсинга одной и той же про-иРНК исследованы для обезьяньего вируса БУ 40.
Активность многих белков определяется их посттрансляционной модификацией — фосфорилированием, ацетилированием, а в ряде случаев расщеплением исходной полипептидной цепи на более мелкие фрагменты.
Широко распространен механизм регуляции активности ферментов, основанный на присоединении к ним молекул-эффекторов. Чаще всего в роли эффекторов выступают конечные продукты цепей биосинтеза, которые связываются с первым или с одним из первых ферментов данного метаболического пути и подавляют его активность, тем самым выключая всю цепь синтеза. Это ингибирование конечным продуктом, благодаря которому регулируются сразу несколько этапов метаболизма. Конечный продукт связывается с ферментом не в его активном центре, а в аллостерическом центре, и такое взаимодействие индуцирует изменение (инактивацию) активного центра фермента.
Таким образом, дифференциальная активность генетического материала может обеспечиваться регуляцией разных уровней его экспрессии: от репликации до ферментативной активности белков, генных продуктов.