Внутриклеточные сигнальные системы

В растениях концентрация цАМФ крайне низка, что объясняется ее превращением в АМФ под действием другого фермента. Однако под действием внешнего раздражителя концентрация цАМФ может увеличиться в течение нескольких секунд в десятки раз, что приводит к активации цАМФ-зависимых протеинкиназ. Это вызывает фосфорилирование разных белков, в том числе и факторов регуляции транскрипции и экспрессии… Читать ещё >
Внутриклеточные сигнальные системы (реферат, курсовая, диплом, контрольная)
В настоящее время говорят о шести сигнальных системах, передающих раздражение, сигналы, в клетке — аденилатциклазной, фосфатидилинозитольной, липоксигеназной, НАДФН-оксидазной (супероксидсинтазной), NO-синтазной и MAP-киназном каскаде.
Аденилатциклазная система названа по имени главного фермента аденилатциклазы, с помощью которой из АТФ образуется циклический АМФ. Аденилатциклаза связана с плазмалеммой. Освобождаемая при гидролизе фосфатных групп энергия может использоваться для синтеза соединений, протекающих с затратой энергии.
Циклический аденозинмопофосфат активирует многие ферменты.
В растениях концентрация цАМФ крайне низка, что объясняется ее превращением в АМФ под действием другого фермента. Однако под действием внешнего раздражителя концентрация цАМФ может увеличиться в течение нескольких секунд в десятки раз, что приводит к активации цАМФ-зависимых протеинкиназ. Это вызывает фосфорилирование разных белков, в том числе и факторов регуляции транскрипции и экспрессии различных генов (см. гл. 2). Установлено, что с работой этой системы связана устойчивость многих растений к различным факторам.
В последнее время появляются доказательства того, что и циклический гуанозинмонофосфат (цГМФ) тоже функционирует в качестве вторичного мессенджера в растениях, подобно животным. В животных клетках цГМФ регулирует ионные каналы и протеинкиназы.
Фосфатидилинозитольная система. Фосфатидилинозитольный цикл был открыт А. Мейблом и Л. Хокиным в животных клетках в 1953 г. (рис. 1.43), однако его роль долгое время была неизвестна. Через много лет оказалось, что образуемые во время функционирования цикла инозитолтрифосфат (ИТФ) и диацилглицерол (ДАГ) являются вторичными мессенджерами. Фосфатидилинозитол — один из липидов плазмалеммы, постоянно присутствующий в мембранах клетки, но в очень малых количествах.
Под действием внешнего фактора изменяется структура молекулы белка-рецептора (Р), находящегося в плазмалемме (см. рис. 1.43). Это изменение конформации рецептора передается G-белку, находящемуся в мембране рядом, а от него — ферменту фосфолипазе С, которая функционирует на внутренней стороне плазмалеммы. Фосфолипаза С становится активной и катализирует реакции, в результате которых образуются два вторичных мессенджера: диацилглицерол (ДАГ) и ииозитол-1,4,5-трифосфат (ИТФ).
Пока молекула ГТФ остается связанной с молекулой G-белка, он успевает активизировать до 10 молекул фосфолипазы С, входящей в состав мембранных белков. После этого происходит гидролиз ГТФ; ГДФ и фосфат освобождаются, G-белок переходит в неактичное состояние и остается в нем, пока не захватит новую молекулу ГТФ.
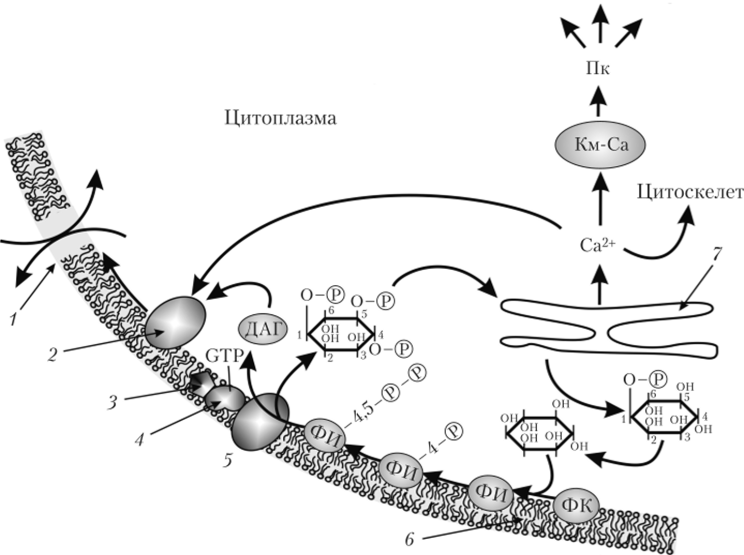
Pwc. 1.43. Фосфатидилинозитольный цикл1:
ДАГ — диацилглицерол; Км-Са — комплекс кальмодулина с кальцием;
Пк — протеинкиназы; 1 — ионные каналы, или насосы; 2 — протеинкиназа С;
- 3 — рецептор; 4 — G-белок; 5 — фосфолипаза С; 6' — плазмалемма;
- 7 — эндоплазматический ретикулум; ФИ — фосфатидилинозитол;
ФИ-4-Р — фосфатидил и нозитол-4-фосфат;
ФИ-4,5-Р-Р — фосфатидилинозитол-4,5-дифосфат; ФК — фосфатидная кислота
Растворимый в воде ИТФ диффундирует в цитозоле до тех пор, пока не встретится с рецепторами на мембране эндоплазматического ретикулума или тонопласта. При взаимодействии рецепторов с ИТФ открываются Са+2-каналы. Поскольку концентрация кальция в цитозоле крайне низка, то он быстро выделяется из ЭПР и вакуоли в цитозоль по градиенту концентрации. Время жизни ИТФ — менее одной секунды, это объясняет очень быстрый выход кальция в цитозоль. В результате концентрация Са+2 в цитозоле увеличивается примерно с 1 до 100 ммоль/л. Ионы кальция начинают поступать в цитозоль через 2 мин после получения сигнала. Затем ИТФ постепенно гидролизуется до инозитола. Далее инозитол включается в состав фосфатидилинозитола и только в составе мембраны фосфорилируется до фосфатидилдиинозитолдифосфата и используется еще раз.
Одновременно с ИТФ ДАГ при участии Са+2 активирует находящуюся в плазмалемме протеинкиназу С. Активированная протеинкиназа С фосфорилирует белки кальциевых каналов плазмалеммы и таким образом 1
Цит. по: Полевой В. ВСаламатова Т. С. Физиология роста и развития растений. Л.: Изд-во ЛГУ, 1991.
активирует их работу. Увеличение концентрации Са2+, в свою очередь, активирует протеинкиназы и другие ферменты, о чем мы уже говорили.
Фосфатидилинозитольный цикл играет важную роль в растении. Например, он участвует в передаче сигнала о водном дефиците, что вызывает закрывание устьиц. Фосфатидилинозитолдифосфат является важным модулятором при взаимодействии мембран с цитоскелетом. Так, повышение его доли в мембране вызывает сборку актиновых филаментов, а дефосфорилирование приводит к разборке цитоскелета.
Липоксигеназная система получила свое название от имени фермента липоксигепазы. Для мембран клеток растений, по сравнению с животными клетками, характерно высокое содержание ненасыщенных жирных кислот — линолевой и линоленовой. Липоксигеназа окисляет эти полиненасыщенные жирные кислоты, присоединяя кислород по месту двойных связей. Субстратом для нее могут быть как свободные полиненасыщенные жирные кислоты, так и находящиеся в составе мембранных липидов.
В липоксигеназной системе участвует и другой фермент — фосфолипаза А. Она активируется кальцием, а также многими биогенными и абиогенными факторами. После ее активации начинается гидролиз мембранных фосфолипидов, и появляются свободные линолевая и лииоленовая кислоты. Кроме того, кальций активирует липоксигеназу. Освободившаяся из липидов линолевая кислота под действием липоксигепазы превращается в нерекисное соединение и далее участвует в образовании жасминовой кислоты (см. параграф 7.2). Жасминовая кислота блокирует Са2+-каналы, что предотвращает избыточное поступление кальция. Затем жасминовая кислота может метилироваться, превращаясь в физиологически активный метилжасмонат, или присоединяет моносахарид и переходит в неактивную запасную форму.
У арабидопсиса обнаружены два гена, ответственные за синтез липоксигеназы. Один из них экспрессируется в листьях, корнях, проростках при нормальном развитии и не зависит от внешних условий. Экспрессия другого гена повышается в ответ на механические повреждения и инфекцию.
Активность липоксигеназ может повышаться двумя способами:
- 1) активацией имеющихся;
- 2) синтезом новых молекул в результате индукции экспрессии генов.
Сильную активацию липоксигеназного пути вызывают высокая температура, водный стресс, ультрафиолетовые лучи, действие озона, ионы кальция, перекись водорода, обезвоживание, действие некоторых гормонов, инфекция.
НАДФН-оксидазная система (супероксидсинтазная) сигнальная система. В этой сигнальной системе участвуют активные формы кислорода (АФК): супероксидный анион (02), гидроксильный радикал (ОН) и перекись водорода (Н202) (см. подпараграф 4.4.2). АФК вызывают перекисное окисление ненасыщенных жирных кислот, окисление SH-rpynn в белках, повреждение ДНК, деполимеризацию некоторых полисахаридов и другие реакции.
Возникновение активных форм кислорода является одним из самых ранних ответов растительной клетки на любое повреждение, в том числе на инфекцию. В нормально функционирующей клетке существует баланс между активацией и дезактивацией кислорода, поэтому количество АФК остается на безопасном уровне. Однако нарушение структуры и функций клетки вызывает, как правило, интенсивную генерацию АФК.
Предполагают, что при окислении молекулярным кислородом НАДФН, находящегося в плазмалемме, образуется супероксид анион, который при участии фермента супероксиддисмутазы (СОД) превращается в перекись водорода. Супероксид-анион и перекись водорода являются вторичными мессенджерами в данной системе. Перекись водорода вызывает активацию факторов регуляции транскрипции и, как следствие, экспрессию генов, в том числе и защитных.
Важную роль в этой сигнальной системе играет салициловая кислота (см. гл. 10). Салициловая кислота связывает каталазу, разлагающую перекись водорода, поэтому количество последней увеличивается. Гибель клеток коррелирует с содержанием салициловой кислоты. Имеются данные о существовании протеинкиназ, активируемых салицилатами, что может объяснять экспрессию защитных генов как салициловой кислотой, так и перекисью водорода.
NO-синтазная сигнальная система связана с образованием оксида азота (NO). Он получается при окислении аргинина. Реакция катализируется синтазой оксида азота, находящейся в цитозоле:
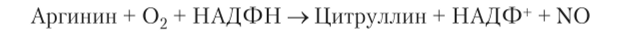
Кроме того, NO может образоваться с помощью нитратредукгазы из нитрита:
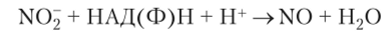
В настоящее время неизвестно, какой из этих двух путей важнее при синтезе оксида азота и как он регулируется.
У растений оксид азота — важный вторичный мессенджер при передаче сигналов и в защитных реакциях. Он стимулирует выход кальция из внутриклеточных депо. Увеличение концентрации кальция в цитозоле активирует сигнальные каскады. NO индуцирует открывание устьиц, вместе с АБК регулируя ширину устьичной щели (см. параграф 8.2), участвует в запуске программированной клеточной смерти и образовании фитоалексинов. Он также индуцирует синтез салициловой кислоты.
MAP-киназный каскад. В клетке существует серия протеинкиназ, которые последовательно фосфорилируют друг друга, в результате происходит каскад реакций (от фр. cascade — многоступенчатый процесс). Этот каскад был назван МАРК-каскадом по имени митогенактивируемой протеинкиназы (англ. МАРК — Mitogen Activated Protein Kinase). Митогеп — это общее название для веществ, которые стимулируют митоз и другие реакции. MAP-киназный каскад — это цепь реакций, в которой одни протеинкиназы регулируются через фосфорилирование другими протеинкиназами.
Первая киназа в этой последовательности ферментов называется МАРККК (МАР-киназа-киназа киназы), ее активирует G-белок. Именно эта киназа запускает весь МАРК-каскад. Она передает фосфорный остаток другой киназе — МАРКК (MAP-киназа киназы), которая, в свою очередь, передает сигнал МЛР-киназе (МАРК). Этот МАРК-каскад является важным способом передачи внутриклеточного сигнала у растений.
Биологический смысл этой цепи реакций состоит в усилении первичного сигнала. MAP-киназа играет главную роль в каскаде: в дальнейшем она фосфорилирует другие белки. Например, она направляется к ядру, где активирует другие протеинкиназы и различные транскрипционные факторы, которые связываются и вызывают дифференциальную экспрессию генома. MAP-киназный каскад выполняет важную регуляторную функцию в процессе развития и дифференцировки клетки, участвует в системе защиты от патогенов. Эта система включается в ответ на патогены и абиотический стресс.
Не всегда реакция клетки на раздражение может индуцироваться только одной сигнальной системой, иногда для этого необходимо взаимодействие нескольких сигнальных систем. Имеется много фактов об активации или инактивации одних сигнальных систем вторичными мессенджерами других. Например, NO-синтазная система подавляет некоторые сигнальные системы.
Несмотря на огромное количество первичных сигналов (раздражений), при ответе клетка часто активизирует одни и те же компоненты сигнальных цепей (это так называемое явление cross-talk). Специфический ответ клетки предполагает, что 1) разные клетки организма отвечают на один и тот же сигнал по-разному; 2) одна и та же клетка отвечает на разные сигналы по-разному. Восприятие сигнала зависит от состояния, в котором находилась клетка перед его воздействием, т. е. от набора рецепторов, наличия вторичных мессенджеров и ир.
Таким образом, в клетках существует скоординированная сигнальная цепь, в которой одни сигнальные системы активируют или инактивируют другие сигнальные системы. Интересно, что аденилатциклазная и Са-фосфоинозитольная сигнальные системы являются эволюционно более древними, чем липоксигеназная, супероксидсинтазная и NO-синтазиая системы, поскольку первые не используют для своего функционирования молекулярный кислород, тогда как вторые не могут функционировать в отсутствие кислорода.