Билатеральные механизмы обучения

Таким образом, в динамике выработки условного рефлекса поздний позитивный потенциал вначале появляется в контралатеральном полушарии, затем в ипсилатеральном, что в итоге приводит к однотипности характера активности обоих полушарий, а на пятом этапе он вызывается только в контралатеральном. На пятом этапе число сигнальных вспышек, сопровождающихся поздними позитивными колебаниями, уменьшается… Читать ещё >
Билатеральные механизмы обучения (реферат, курсовая, диплом, контрольная)
Билатеральность обучения, то есть возможность переноса результатов обучения с одного полушария на другое, была впервые установлена в лаборатории И. П. Павлова его учениками: Красногорским (1911), Быковым (1924).
Несколько позже появились работы, освещающие некоторые механизмы межполушарного переноса положительных временных связей. Так, К. С. Абуладзе (1953, 1961) показано, что его наличие или отсутствие зависит от интенсивности подкрепляющего стимула. Длительное упрочение одностороннего рефлекса приводит к исчезновению феномена переноса (Абуладзе и др., 1976).
В работах Н. И. Лагутиной (1951) и Н. И. Николаевой (1955) было показано, что при выработке условного рефлекса с использованием в качестве условного сигнала прямого раздражения коры или подкорковых структур, перенос сигнального раздражения на симметричную раздражаемой структуру вызывает условную реакцию.
Межполушарный перенос обучения происходит при выработке положительных условнорефлекторных связей. При угашении рефлекса перенос стимула на противоположную раздражаемой сторону требует повторного угашения условнорефлекторной реакции, то есть тормозные связи не переносятся с одного полушария на другое.
Билатеральность обучения показана на различных видах животных. Причем в эволюционном ряду межполушарный перенос обучения обеспечивается комиссурами, которые у данного вида животных осуществляют высшие интегративные функции. У рыб это текто-мозжечковые структуры, у амфибий — структуры среднего мозга, у птиц — полосатые тела, а у млекопитающих — кора головного мозга.
У кошек решающим фактором переноса обучения в пределах зрительного анализатора является передача информации по волокнам мозолистого тела и передней комиссуре (Myers, 1955, 1965).
В основе межполушарного переноса обучения лежит обмен информацией между полушариями через мозолистое тело и другие спайки мозга.
Доти и соавторы (Doty et. al., 1974) решающее значение в межполушарном переносе обучения отводят передней комиссуре. Т. С. Наумова отводит мозолистому телу роль совершенствования межполушарных отношений, поскольку именно с ним связаны тормозные влияния на симметричные пункты коры, которые вначале облегчают образование рефлекса, а затем препятствуют участию одного из полушарий в его реализации. Правомерность такого представления подтверждают исследования В. Л. Бианки (1976), в которых показано расширение фокуса максимальной активности на световые стимулы в зрительной коре мозга при перерезке мозолистого тела. Следовательно, в норме межполушарные связи через мозолистое тело препятствуют иррадиации возбуждения по коре и повышают дифференцируемость корковой мозаики при условнорефлекторной деятельности.
Участие мозолистого тела в переносе энграммы памяти из обученного полушария в необученное подтверждают данные об улучшении свойств вторичного следа, формируемого в симметричной обучаемой корковой зоне, под действием пирацетама, специфически облегчающего каллозальные ответы (Й. Буреш, О. Бурешова, 1960).
В наших экспериментах по электрофизиологическим и поведенческим коррелятам прослежены отношения симметричных систем коры мозга в исходном состоянии и их качественные перестройки в ходе выработки условных рефлексов.
Уже по показателями поведенческих условно-рефлекторных двигательных оборонительных реакций на сигнал стало ясно, что межполушарное взаимодействие в процессе формирования рефлекса характеризуется сменой функциональных состояний симметрии и асимметрии. Внешне это проявилось в определенной динамике двигательных реакций на сигнал, что позволило нам выделить 5 этапов взаимодействия полушарий головного мозга при выработке оборонительного условного рефлекса.
На первом этапе сигнал еще не вызывает условной двигательной реакции, двигательный ответ развивается только на подкрепление. На втором этапе на стороне подкрепления появляются локальные двигательные реакции на сигнал. Видимо, на этом этапе в организации условного рефлекса доминирующую роль играет то полушарие мозга, в которое преимущественно адресовалось подкрепляющее раздражение. На этом этапе генерализация условного рефлекса еще не охватывала симметричные пункты второго полушария. На 3-м и 4-м этапах двигательные реакции на сигнал были двусторонними, но на 3-м этапе они были синергичными, а на 4-м — реципрокными. Различия в характере эффекторных реакций на 3-м и 4-м этапах, видимо, обусловлены тем, что на 3-м этапе выработки рефлекса возбуждение одновременно охватывает моторные центры полушарий, обеспечивающие флексорную реакцию. Это приводит к торможению, на основе индукционных отношений, центров, обеспечивающих экстензию конечности. На 4-м этапе, вследствие усиливающегося тормозного влияния со стороны доминирующего полушария, флексорный центр в субдоминантном полушарии тормозится, а это приводит к временному возбуждению экстензорного центра, что и проявляется реципрокностью двигательных реакций на сигнал.
На пятом этапе выработки рефлекса сигнал вызывал локальные условные реакции только на стороне подкрепления. Следовательно, на этом этапе генерализация сменялась концентрацией нервных процессов в полушарии, организующем условно-рефлекторный ответ. Такая динамика наблюдалась при выработке оборонительного условного рефлекса с применением в качестве подкрепления электрокожного раздражения лапы или уха животного.
Выявленная в поведенческих опытах динамика межполушарных отношений при формировании условно-рефлекторных двигательных реакций ставят вопрос о существовании различий полушарной активности априорно, т. е. в фоновом и вызванном режимах.
У большинства животных в субдоминантном полушарии было больше нейронов с пачечной и групповой формами активности, а в доминантном с тонической. Средняя частота импульсных разрядов была больше у нейронов, зарегистрированных в коре доминирующего полушария. Кроме того, нейронная активность доминирующего полушария имела большую вариабельность и меньшую скоррелированность импульсных потоков соседних нейронов.
Выявленные отличия в наших исследованиях по фоновой активности доминирующего и субдоминантного полушарий подтвердились исследованиями с применением световой и электрокожной стимуляций.
Межполушарные отличия однотипных зон коры отмечены и при анализе фокальной активности. Световое раздражение вызывало в 17-м поле доминирующего полушария достоверно более коротколатентные (сравнительно с субдоминантным) фокальные ответы. В 18-м поле, наоборот, коротколатентные ответы были в субдоминантном полушарии. Неадекватные стимулы, хотя и вызывали изменения фокальной активности коры, но они были однотипны в обоих полушариях.
В 17-м поле зрительной и соматосенсорной коры доминантного полушария реагирующих нейронов было больше (за счет возбуждающихся), латентные периоды их реакций были короче, вариабельность средней частоты потоков импульсов ниже, чем в субдоминантном. В 18-м поле зрительной коры превосходящее число реагирующих нейронов отмечено в субдоминантном полушарии. В нем же была несколько большая средняя частота разрядов нейронов, короче ЛП реакций, однако, вариабельность средней частоты была выше в доминантном полушарии.
В 17-м и 18-м полях зрительной коры количество полисенсорных нейронов превалировало в доминантном полушарии.
При неадекватной стимуляции нейрональная активность симметричных пунктов коры, как правило, мало отличалась. В этом, видимо, проявляется эволюционное развитие анализаторных областей коры, заключающееся в разделении ее на зоны с преобладанием элементов, настроенных реагировать на специфические раздражители (моносенсорных нейронов), и зоны с преобладанием элементов ассоциативного типа (полисенсорных нейронов). Одновременно с этим происходит дифференциация функций между полушариями.
Нашими опытами установлено, что функциональное доминирование возникает лишь в случае, если применяется раздражение, адекватное по своей сенсорности для коркового конца данного анализатора. Так, применение светового раздражения сопровождалось межполушарной асимметрией зрительных зон коры, а при применении электрокожного раздражителя отмечалось состояние симметрии.
В динамике образования оборонительного условного рефлекса, что прослеживалось по изменениям суммарной, фокальной и импульсной активности межполушарные различия этих показателей были существенно иными, чем при действии индифферентных стимулов. Это проявилось в том, что в динамике образования условного рефлекса корковые процессы имеют специфические особенности на разных этапах его формирования.
Как уже отмечалось выше, на основании анализа опытов с выработкой оборонительного и пищевого рефлексов нами было выделено пять этапов становления условного рефлекса. Эти этапы определялись не по характеру электрической активности, а по динамике условной двигательной реакции. Однако, упрочение условного рефлекса имело на этих этапах и специфические особенности, отражаемые в активности корковых процессов.
На первом этапе, когда двигательная реакция возникала только на безусловный стимул, отличий в суммарной биоэлектрической активности верхних и нижних слоев симметричных зон коры не было (рис. 18, 1 этап обучения).
На втором этапе появилась реакция усвоения ритма условной стимуляции в верхних слоях полушария, контралатерального по отношению к подкрепляющему стимулу (рис. 18 11 этап обучения).
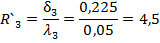
Рис. 18. Динамика электрокортикограммы верхних и нижних слоев симметричных пунктов зрительной коры мозга кошки на разных этапах выработки условного рефлекса. Внизу: отметка условного и безусловного раздражителей.
На третьем этапе условная реакция усвоения ритма стимуляции охватывала все слои контралатерального и верхние слои ипсилатерального по отношению к подкреплению полушария (рис.18 111 этап обучения).
На четвертом этапе условная реакция усвоения ритма стимуляции охватывала все слои симметричных зон обоих полушарий (рис. 18 IV этап обучения). На этом этапе происходило замыкание рефлекса и появлялись условные двигательные реакции.
На пятом этапе при упроченном рефлексе условная реакция усвоения ритма стимуляции локализовалась в нижних слоях полушария, контралатерального по отношению к подкрепляющему стимулу Рис. 18. V этап обучения).
Таким образом, анализ суммарной активности показал, что усвоение ритма в динамике выработки рефлекса вначале наиболее выражено в верхних слоях коры контралатерального относительно стороны подкрепления полушария, а затем в нижних слоях контралатерального относительно стороны подкрепления полушария, верхних слоях контралатерального, и, наконец, нижних слоях контралатерального полушария. При упроченном рефлексе наибольшие изменения ЭкоГ-активности сохраняются в глубоких слоях коры контралатерального полушария. Такая динамика имела место в разные периоды: в предсигнальный, сигнальный, подкрепления и последействия. Однако, время появления таких изменений от начала выработки рефлекса для всех периодов не было одинаковым.
Раньше всего они возникали в последействии, затем в периоды подкрепления и действия сигнала и, наконец, в предсигнальный. Следовательно, начальные изменения активности происходят на следах предшествующих сочетаний и это характерно для обоих полушарий Исследования, проведенные с применением регистрации фокального потенциала, принципиально подтвердили динамику развития межполушарного взаимодействия, выявленную по ЭкоГ-показателям, но при этом определились некоторые особенности динамики возбудительных и тормозных процессов. В качестве критерия выработки рефлекса нами было использовано появление в фокальном ответе позднего позитивного колебания (Рис.19).
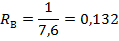
Рис. 19. Фокальные вызванные потенциалы симметричных пунктов поля 18 зрительной коры. И — ипси-, К — контралатеральное полушарие относительно стороны подкрепляющего раздражения. Отметка времени — 50 мс.
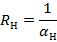
Рис. 20. Фокальные ответы симметричных пунктов поля 18 зрительной коры. Римские цифры — этапы выработки условного рефлекса. Стрелки — момент предъявления вспышки света. Верхние осциллограммы в каждой паре — запись из коры контралате-рального полушария, нижние — ипсилатерального. Отметка времени — 50 мс.
На первом этапе выработки рефлекса в фокальных ответах поздний позитивный потенциал на вспышки света, используемые как сигнал, не встречался. Первичные негативные колебания в разных полушариях имели однотипные величины латентных периодов и длительностей. По мере упрочения рефлекса появлялись различия в характере ответов, регистрируемых в разных полушариях: в ипсилатеральном — длительность первичных негативных колебаний, латентные периоды и их вариабельность становились больше (рис.20).
Поздний позитивный потенциал, появление которого свидетельствует об усилении тормозных процессов, на первых этапах выработки рефлекса был мало отличим в обоих полушариях, но количественно в ответах на повторяющиеся вспышки света он преобладал в полушарии, контралатеральном относительно подкрепления. Следовательно, тормозные явления в последнем были более интенсивны.
Начиная с третьего этапа, межполушарные различия проявлялись по характеру поздних позитивных колебаний. В ипсилатеральном полушарии ЛП и длительность позитивных волн были больше, чем в контралатеральном. Следовательно, лабильность нервных процессов в последнем была выше.
Повторение сочетаний на четвертом и пятом этапах приводило к резкому урежению появления поздних позитивных колебаний в фокальных ответах, регистрируемых из ипсилатерального полушария. Скорость появления этих колебаний в контралатеральном полушарии и урежение их в ипсилатеральном были выше, если латеральность подкрепления совпадала с доминирующей по лапе стороной, и наоборот.
В динамике выработки рефлекса изменялся рисунок позднего позитивного колебания: передний фронт его удлинялся, задний — укорачивался. Эти изменения в основном отмечались в полушарии, контралатеральном относительно подкрепления.
Таким образом, в динамике выработки условного рефлекса поздний позитивный потенциал вначале появляется в контралатеральном полушарии, затем в ипсилатеральном, что в итоге приводит к однотипности характера активности обоих полушарий, а на пятом этапе он вызывается только в контралатеральном. На пятом этапе число сигнальных вспышек, сопровождающихся поздними позитивными колебаниями, уменьшается. Так как эти колебания свидетельствуют об усилении тормозных процессов, то, следовательно, по мере выработки рефлекса торможение вначале растет в контралатеральном, а затем ипсилатеральном полушариях; при упроченном же рефлексе оно более выражено в контралатеральном.
Анализ нейронной активности, регистрируемой в симметричных пунктах зрительной и соматосенсорной коры, показал, что условно-рефлекторное обучение сопровождается определенным изменением последовательности разрядов отдельных нейронов. Эти изменения в разных полушариях были выражены неоднозначно.
Хорошо известно, что в динамике выработки условного рефлекса уменьшается или полностью исчезает условная активность нейронов сигнальной зоны коры Шульговский, Котляр, 1968. Однако, исчезновение реакции нейронов в форме изменения частоты разрядов (признак, на который опирается большинство авторов) не может служить доказательством прекращения участия нейронов в условно-рефлекторном процессе.
В работах Doty 1974, Э. Г. Заркешева 1980, 1982 указывается, что в любой структуре мозга имеются обучающиеся клетки, реализующие замыкание данного вида временной связи. В нашем исследовании нейроны, усиливающие реакцию на сигнал в начале угашения рефлекса были отнесены к «обучающимся», а не изменявшие — к «необучающимся». У «обучающихся» нейронов в динамике выработки условного рефлекса независимо от зоны регистрации вариабельность частоты разрядов уменьшалась в фоне при предъявлении сочетаний и в последействии. Параллельно этому увеличивалась корреляция потоков импульсных разрядов нейронов, зарегистрированных одновременно по одному каналу.
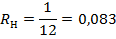
Рис. 20. Импульсная активность пары нейронов симметричных пунктов зрительной коры. Верхние нейронограммы — из ипсилатерального, нижние — из контралатерального относительно стороны подкрепления полушария. Цифры слева — номера сочетаний. Стрелка — момент предъявления сигнальной вспышки света. Отметка времени — 50 мс.
«Обучающиеся» нейроны реагировали на сигнальные стимулы с более длинными латентными периодами. В структуре их импульсного потока (в динамике выработки условного рефлекса), как правило, формировалась тормозная пауза, которая по времени строго соответствовала позитивному колебанию фокального ответа, регистрируемого тем же электродом, что и импульсный разряд. если для «необучающихся» нейронов было характерно изменение степени активности — вначале усиление с последующим снижением ее до исходного уровня, то у большинства «обучающихся» на первых этапах выработки рефлекса частота разрядов в ответах на сигнал также увеличивалась, а затем их разряды приобретали особый порядок следования импульсов Рис. 20.
Как уже отмечалось, исследования нейрональных механизмов обучения по электрическим проявлениям показали, что по мере становления и упрочения рефлекса реакции фоновой ритмики, а также вызванные потенциалы и импульсная активность, сопровождающие условно-рефлекторную реакцию, ярко выраженные вначале, на более поздних стадиях ослабевают или не выявляются вовсе Н. Н. Василевский, 1968. Это может быть связано как с реализацией выработанного условного рефлекса лишь небольшим числом клеточных элементов, так и обеспечением условно-рефлекторных реакций скорее перестройками узора импульсного потока, чем существенными изменениями его интенсивности.
Исследования в опытах с оборонительным условным рефлексом, динамики его становления, позволили нам предположить, что значительная часть нейронов для организации условной реакции вначале использует частотный код, который по мере упрочения условного рефлекса не выявляется. Однако, анализ узора потока межимпульсных интервалов позволил заметить стабильное увеличение частоты встречаемости некоторых из них — определенных для данного нейрона.
Наши данные показывают, что нейроны с такими формами активности встречаются в фоне в относительно малом числе случаев, но в динамике выработки рефлекса встречаемость таких нейронов возрастала. Причем больше таких нейронов было в 18 поле, сравнительно с 17 полем, зрительной коры. Следовательно, пачечная и групповая форма активности нейронов являются одними из форм кодирования сигнальной информации при условно-рефлекторной деятельности. Тем более, что, по мнению В. Л. Силакова 85, переход на пачечный режим обеспечивает высокую надежность передачи информации, поскольку эффективность пачки импульсов выше, а вероятность случайной блокады ее ниже, чем одиночных разрядов.
При выработке рефлекса повышается сходство активности по характеру межимпульсных интервалов в фоне с интервалами в момент условной реакции. Это свидетельствует, что в нейрональной системе происходят существенные перестройки на оптимальное решение задачи до предъявления пускового сигнала Сравнение динамики межимпульсных интервалов потоков активности нейронов симметричных пунктов коры показало, что при выработке оборонительного условного рефлекса перестройки характера импульсной активности раньше происходят в коре контралатерального полушария, затем появляются в коре ипсилатерального. На этапе упроченного рефлекса, когда имела место изолированная односторонняя двигательная реакция, в ответ на сигнал специфический узор возникал у нейронов только этого полушария.
Видимо, в обоих полушариях способы кодирования сигнальной информации одинаковы, но на стадии упроченного рефлекса световое раздражение, используемое при выработке оборонительного условного рефлекса, в ипсилатеральном полушарии не воспринимается как сигнальное. В этом, видимо, проявляются взаиморегулирующие отношения полушарий головного мозга при условно-рефлекторной деятельности.
Таким образом, как и в опытах с регистрацией суммарной, фокальной активности, так и опытах с регистрацией импульсной активности отмечалась единая динамика межполушарных отношений: в начале выработки рефлекса стирались существующие в фоне полушарные особенности активности коры, затем усиливалось сходство ее по всем исследуемым зонам, и, наконец, когда имела место локальная односторонняя двигательная реакция на сигнал, формировалась новая функциональная межполушарная асимметрия.
Условно-рефлекторные эксперименты позволяют заключить, что у животных помимо врожденной межполушарной асимметрии функций, возникает функциональная асимметрия и в результате обучения. В этом плане мозг человека не является исключением.