Биологический вид.
Особенности расовых различий

Действие отбора придает указанным особенностям популяций черты целесообразности. Благодаря отбору одновременно с приспособленностью к существующим условиям популяции приобретают также определенную степень приспособляемости, т. е. способность выдержать разного рода изменения условий. Приспособляемость популяций определяется, с одной стороны, широтой нормы реакции составляющих популяцию генотипов… Читать ещё >
Биологический вид. Особенности расовых различий (реферат, курсовая, диплом, контрольная)
Прежде чем мы приступим к анализу основных факторов, определяющих изменения генофонда вида, необходимо разобраться, что представляют собой основные структурные единицы эволюционного процесса — биологический вид и его популяции. Как мы уже упоминали, начиная с работ Ч. Дарвина, биологический вид оказался в центре внимания эволюционистов. Не будет преувеличением сказать, что синтетическая теория эволюции представляет собой главным образом теорию видообразования. Проблема видообразования находится в центре внимания и некоторых альтернативных эволюционных концепций (сальтационизм, пунктуализм). При этом представления о биологическом виде претерпели со времени Ч. Дарвина значительные изменения.
Вид как биологическая макросистема Подавляющее большинство организмов в той или иной форме используют половой процесс как важнейший способ обмена генетической информацией между разными особями. Половой процесс не обязательно связан с размножением (например, конъюгация инфузорий или меромиксис бактерий), но всегда объединяет различных особей в единую систему — биологический вид. (Меромиксис — половой процесс у бактерий, при котором происходит конъюгация двух бактериальных клеток и часть хромосомы одной из них переходит в другую). Благодаря половому процессу и мейозу в пределах вида постоянно возникают новые комбинации наследственных признаков, новые генотипы, которые все вместе составляют генофонд вида как целого. Биологический вид, состоящий из многочисленных особей, генетически родственных, но различающихся по своим комбинациям наследственных признаков, представляет собой целостную биологическую макросистему.
Поскольку вид является стохастической (вероятностной) системой, характеризующейся нежесткими связями составляющих его компонентов — особей, его устойчивость к изменениям внешней среды определяется в каждом конкретном случае наиболее удачными комбинациями признаков в генотипах отдельных особей. Этим вид отличается от жестких (корреляционных) систем, подобных отдельным организмам, устойчивость которых определяется наименее устойчивыми в данных условиях компонентами. Так, к гибели данного организма ведет летальная мутация в каком-то одном локусе, т. е. изменение одного гена, тогда как для вида в целом гибель отдельных особей, оказавшихся хуже приспособленными в данных условиях или погибших от случайных причин, является относительно малосущественным фактором. Высокая устойчивость биологического вида к изменениям внешней среды определила его роль как основной формы организации живой материи.
Вид, объединяемый в единое целое с помощью полового процесса, неизмеримо устойчивее к изменениям среды, чем клон — потомство данной особи, полученное посредством бесполого размножения. К организмам, размножающимся исключительно бесполым способом (деление, почкование и т. п.), понятие биологического вида, в сущности, неприменимо. Систематики для удобства описаний таких форм иногда используют термин «агамный вид». Аналогично формы организмов, размножающиеся исключительно путем партеногенеза, называют партеновидами.
В предыдущей главе мы уже рассматривали значение разных форм изменчивости для выживания вида, подчеркнув особую роль резерва комбинативной изменчивости, пополняемого новыми мутациями. Благодаря действию естественного отбора генофонд вида представляет собой не случайный набор аллелей, а определенным образом сбалансированную систему, в которой относительное количество тех или иных аллелей определяется приспособительной ценностью контролируемых ими фенотипических особенностей в данных условиях.
На основе генетической общности регулярно скрещивающихся друг с другом особей формируется их морфологическое, физиологическое и поведенческое сходство, на которое прежде всего обращали внимание систематики, выделяя разные виды организмов именно по критериям морфофизиологических (а в большинстве случаев только морфологических) сходств и различий.
Хотя виды организмов представляют собой реальные биологические системы, дать четкое и исчерпывающее определение понятию «биологический вид» оказалось нелегко. Причины этого коренятся в самой природе этой формы организации живой материи.
Долгое время вид определяли как совокупность морфологически и физиологически сходных особей, способных скрещиваться друг с другом, давая плодовитое потомство. Для практических целей такое определение было удобно, поскольку, казалось, давало вполне отчетливые критерии для идентификации разных видов. Однако по мере накопления новых данных систематики встречались с существенными трудностями в реальном применении указанных критериев.
Прежде всего в таком определении совершенно игнорируется сложная внутренняя структура вида. Уже во времена Дарвина было известно, что многие виды организмов включают устойчивые формы, обитающие в разных частях видового ареала (области географического распространения), — подвиды. Подвиды стабильны и достаточно отчетливо различаются по морфофизиологическим критериям, но в зонах контакта скрещиваются друг с другом, давая плодовитое потомство с промежуточным состоянием признаков (так называемые «зоны интерградации» между подвидами). Ч. Дарвин, изучая изменчивость организмов, пришел к выводу, что индивидуальные различия особей по степени своего выражения плавно переходят в различия подвидов, а различия подвидов столь же постепенно переходят в видовые различия. На основании этого Дарвин заключил, что между видами, подвидами и разновидностями нет принципиальных различий. Этот вывод Дарвина, если понимать его буквально, неверен, поскольку он игнорирует особую биологическую роль вида как формы организации живой материи и как целостной эволюционной единицы. При оценке таксономического статуса многих близких форм организмов возникают сложности — считать ли их подвидами одного вида или самостоятельными, хотя и родственными друг другу, видами.
Популяционная структура вида Вид представляет собой в действительности гораздо более сложную систему, чем просто совокупность скрещивающихся сходных друг с другом особей. Он распадается на более мелкие естественные группировки особей — популяции, представляющие население отдельных относительно небольших участков в пределах всей зоны распространения (ареала) данного вида. Внутри каждой популяции имеет место наибольшая степень панмиксии; скрещивание же особей, происходящих из разных популяций, имеет место относительно более редко, и обмен генетической информацией между разными популяциями более ограничен. Это обусловливает определенную самостоятельность генетических процессов, происходящих в разных популяциях одного вида. В результате каждая популяция характеризуется своим специфическим генофондом с присушим только данной популяции соотношением частот встречаемости разных аллелей и с соответствующими особенностями спектра изменчивости. Эти генетические различия популяций могут иметь как случайный, так и неслучайный характер. Последнее свойственно относительно крупным популяциям (порядка 500 особей и более), длительное время существующим в данном географическом районе.
Природные условия в разных частях ареала вида обычно более или менее различны. В результате отбор имеет разное направление для популяций одного вида, населяющих разные районы. Следствием этого является возникновение относительно устойчивых различий генофондов разных популяций. Особенности популяционных генофондов благодаря действию отбора приобретают приспособительный характер: подбор аллелей в конкретном генофонде, обусловливающий специфическую картину комбинативной и модификационной изменчивости в данной популяции, становится оптимальным для условий обитания этой популяции.
Частота встречаемости разных аллелей в популяции определяется частотой прямых и обратных мутаций, давлением отбора, обменом наследственной информацией с другими популяциями в результате эмиграции и иммиграции особей. При относительной устойчивости условий в достаточно большой популяции все указанные процессы приходят к состоянию относительного равновесия, конкретный характер которого определяется, с одной стороны, спецификой условий, а с другой — генетической системой данного вида. В итоге такие достаточно крупные и устойчивые популяции приобретают сбалансированный и оптимизированный отбором генофонд, особенности которого имеют приспособительный характер и обусловливают специфические черты данной популяции (экологические, поведенческие, а во многих случаях и достаточно определенные морфофизиологические показатели).
Различия популяций, населяющих удаленные или относительно изолированные друг от друга области, становятся более отчетливыми в связи с уменьшением обмена генетической информацией между ними. Результатом достаточно продолжительной изоляции является формирование подвидов, под которыми понимаются популяции данного вида, населяющие разные участки видового ареала (т. е. имеющие аллопатрическое распространение) и характеризующиеся устойчивым комплексом морфологических, физиологических и экологических признаков, закрепленных наследственно. Однако подвиды полностью сохраняют скрещиваемость друг с другом, и если контакт между ними вновь расширяется, возникает зона интерградации, в которой в результате гибридизации особи имеют промежуточное состояние признаков. Наличие в пределах вида нескольких устойчиво различающихся друг от друга подвидовых форм обозначается термином политипичность вида.
Если ареалы отдельных подвидов достаточно велики, подвиды распадаются на популяции меньшего масштаба — экологи различают несколько уровней таких территориальных (аллопатрических) группировок. Таким образам, внутри вида существует сложная иерархическая система территориальных популяций, которая является приспособлением к оптимальному использованию всего разнообразия условий в разных районах видового ареала.
Поскольку популяции обладают специфическим генофондом, находящимся под контролем естественного отбора, очевидно, что эти естественные группировки особей должны играть важнейшую роль в эволюционных преобразованиях вида. Все процессы, ведущие к каким бы то ни было изменениям вида — к его разделению на дочерние виды (видообразование) или к направленному изменению всего вида в целом (филетическая эволюция), начинаются на уровне видовых популяций. Эти процессы преобразований популяционных генофондов принято называть микроэволюцией. По определению Н.В. Тимофеева-Ресовского, Н. Н. Воронцова и А. В. Яблокова, популяции представляют собой элементарные структурные единицы эволюционного процесса, а векторизованные (направленные) изменения генофондов популяций — элементарные эволюционные явления.
Вид в пространстве С учетом внутривидовой популяционной организации Э. Майр определил биологический вид как группу фактически или потенциально скрещивающихся естественных популяций, которая физиологически изолирована от других подобных групп (т.е. от других видов). Следует отмстить, что и это определение не является исчерпывающим, оставляя неопределенность в двух отношениях.
Прежде всего, если две популяции в рассматриваемый момент времени разделены каким-либо географическим барьером, непроходимым для данного вида организмов (горный хребет, морской пролив, пустыня, для водных форм — суша и т. п.), очевидно, они не обмениваются генетической информацией, хотя еще могут сохранять потенциальную способность к скрещиванию (реально не используемую). Следует ли относить такие популяции к одному виду?
Эта неопределенность совершенно неизбежна, так как она является отражением действительной ситуации, нередко наблюдаемой в природе. Многие виды организмов имеют разорванный, прерывистый ареал, распадаясь на группы изолированных друг от друга популяций. Можно назвать хотя бы такие классические примеры разорванного ареала, как распространение голубой сороки (Cyanopica cyana), встречающейся на Пиренейском полуострове и на Дальнем Востоке, или ареал вьюна (Misgurnus fossilis), обитающего в пресных водоемах значительной части Европы и Юго-Восточной Азии, но отсутствующего на территории между 70 и 90 восточной долготы. Дальнейшая судьба ныне существующих популяций любого вида организмов неизвестна — пойдет ли их обособление дальше, или же барьеры, разделяющие популяции, нарушатся, и восстановится обмен генами между разными популяциями. В первом случае изолированные популяции могут дать начало новым самостоятельным видам, во втором они вновь объединяются в один вид. Разумеется, последнее вероятнее при значительно меньшей степени разрыва ареала, чем в двух приведенных примерах, если изолированные в данный момент популяции географически не столь удалены друг от друга. Так или иначе судьба отдельных популяций зависит в первую очередь от дальнейших изменений среды, которые, как правило, непредсказуемы. Слияние популяций, обособленных в течение более или менее длительного времени, с другими популяциями вида придает всякому эволюционному стволу своеобразный «сетчатый» характер.
В некоторых случаях, особенно у видов с достаточно большим ареалом, может возникнуть еще более любопытная ситуация, которую хорошо иллюстрирует следующий пример. Большая синица (Parus major) широко распространена в Евразии — от Ирландии до Японии и Индонезии (рис. 11). В пределах этого огромного ареала орнитологи выделяют свыше 30 подвидов большой синицы. Все подвиды, соседствующие друг с другом, легко скрещиваются с образованием зон интерградации. Расселение большой синицы на восток осуществлялось, по-видимому, по двум направлениям: северному (леса умеренного пояса, к северу от зоны пустынь и гор Центральной Азии) и южному (Персия, тропические леса Индии и Индокитая). На Амуре встретились две формы большой синицы, проникшие сюда по северному и южному путям расселения: подвид Parus major major (распространенный от Западной Европы до Дальнего Востока) и подвид P. major minor. Эти две формы не скрещиваются друг с другом, т. е. ведут себя как самостоятельные виды. Однако P. m. minor легко скрещивается с более Южными подвидами большой синицы, а те, в свою очередь, — с подвидами, обитающими в Индии и Персии; последние формы легко скрещиваются с европейскими P. m. major. Как же приложить определение биологического вида к этой причудливой картине родственных форм? С одной стороны, P. m. major и Р. m. Minor по их нескрещиваемости друг с другом можно оценить как два разных вида; с другой стороны, они связаны через цепь легко скрещивающихся южных популяций, по которым в принципе возможна передача генетической информации от одного конца цепи до другого. Подобные ситуации встречаются не столь уж редко, как можно было бы подумать. Б. Ренш предложил обозначать такую цепь родственных аллопатрических форм термином «круг рас», а если некоторые формы в этом «круге» заслуживают выделения в качестве самостоятельных видов — «кольцевой вид». Позднее Э. Майр назвал такие аллопатрические родственные формы, которые на основании имеющихся данных трудно определить как подвиды или самостоятельные виды, «полувидами», а группу родственных полувидов (т.е. кольцевой вид Б. Ренша) — «надвидом» .
Вторая неопределенность в концепции биологического вида выявляется при попытке приложить определение Майра к популяциям, разделенным не географически, а во времени, т. е. к популяциям, связанным друг с другом родством не по горизонтали, а по вертикали, как предки с потомками. За истекший промежуток времени вид может существенно измениться. Так как изменения генофонда постепенны, невозможно провести отчетливую границу между предковым видом и любым из видов-потомков, если известен весь эволюционный ствол. Эта проблема неизбежно встает перед палеонтологами при изучении групп организмов, Достаточно полно представленных в палеонтологической летописи. Выход можно отчасти найти, используя ряд дополнительных терминов. Непрерывный ряд последовательных во времени популяций, каждая из которых является потомком предшествующей и предком последующей, называют филетической линией. Случайные отрезки этой последней, обычно известные палеонтологам, представляют собой палеонтологические виды (см. рис. 10). Наконец, биологический вид — это отдельная филетическая линия, рассматриваемая в данный момент времени, включая все живущие одновременно популяции, соответствующие определению Э. Майра. Биологический вид является, так сказать, «срезом» филетической линии плоскостью времени, или филетической линией, рассматриваемой в течение кванта времени (А. Кейн).
Д. Симпсон предложил для наименования отдельной филетической линии особый термин «эволюционный вид», дополнив определение филетической линии указанием на ее эволюционную независимость от других линий, собственную эволюционную роль и определенную тенденцию эволюционных изменений. Эта концепция подчеркивает эволюционную целостность вида и может быть использована в анализе филетических линий, переплетающихся друг с другом или расходящихся как независимые ветви.
Приспособленность и приспособляемость вида Как уже отмечалось, внутривидовая популяционная организация является приспособлением к оптимальному использованию всего разнообразия условий в пределах видового ареала. Приспособленность всякой популяции к конкретным условиям ее обитания определяется специфическим подбором аллелей в ее генофонде и в результате этого — соответствующей нормой реакции особей, обусловливающей определенные фенотипические особенности. Различия популяций выражаются прежде всего в характере поведенческих реакций особей, совокупность которых определяет такие важные экологические характеристики популяции, как особенности питания, плотность населения, возрастная и половая структура, сезонные реакции, динамика численности и др.
Действие отбора придает указанным особенностям популяций черты целесообразности. Благодаря отбору одновременно с приспособленностью к существующим условиям популяции приобретают также определенную степень приспособляемости, т. е. способность выдержать разного рода изменения условий. Приспособляемость популяций определяется, с одной стороны, широтой нормы реакции составляющих популяцию генотипов (т.е. возможностями их модификационной изменчивости), с другой же стороны — генетической гетерогенностью, разнообразием аллелей в пределах популяционного генофонда (т.е. запасом комбинативной изменчивости). Важность этого мобилизационного резерва изменчивости для выживания популяций и эволюционную необходимость высокой степени генетической гетерогенности природных популяций показал С. С. Четвериков. При изменениях условий, выходящих за рамки нормы реакции генотипов, которые были оптимально приспособлены к прежним условиям, популяция сможет сохраниться только при наличии в ее генофонде таких генотипов, которые окажутся жизнеспособными и в новых условиях. Следовательно, отбор благоприятствует поддержанию генетической гетерогенности популяций. Запас комбинативной изменчивости включает наряду с оптимальными в данных условиях аллелями также относительно слабо приспособленные аллели. Последние снижают общую приспособленность популяции и составляют так называемый генетический груз, который, по образному выражению Д. Холдейна, является той ценой, которую вынуждена платить популяция за право эволюционировать и, добавим, чтобы выжить и сохраниться во времени. Благодаря этому приспособляемость популяции, которой благоприятствует генетическая гетерогенность и, как следствие, генетический груз, до известной степени противостоит приспособленности популяции к существующим условиям, основанной на оптимальных аллелях. Таким образом, в природных популяциях должно существовать некоторое равновесие между этими двумя противоположными тенденциями.
По определению Т. Мёллера, генетический груз представляет собой ту часть популяционного генофонда, которая определяет появление менее приспособленных особей, подвергающихся избирательной выбраковке (элиминации) под воздействием естественного отбора. Общий генетический груз складывается из нескольких составляющих: 1) мутационный груз — неудачные мутации, существенно снижающие приспособленность; 2) сегрегационный груз — выщепляющиеся в каждом поколении слабо приспособленные гомозиготные особи по таким аллелям, которые в гетерозиготном состоянии обладают высокой приспособленностью (так называемое «сверхдоминирование», см. ниже); 3) субституционный груз — результат изменений приспособленности фенотипических признаков при изменениях внешних условий или появлении новых, более удачных фенотипических вариантов.
Эволюционные преобразования вида под воздействием отбора Неизбежно сопровождаются гибелью хуже приспособленных особей. Общее число отбракованных отбором особей при замещении одного аллеля другим Д. Холдейн назвал «платой за отбор». По Расчетам Холдейна, плата за отбор при одновременном эволюционном замещении нескольких независимых аллелей (разных генов) оказывается чрезвычайно высокой: она экспоненциально возрастает с увеличением числа независимых генов, подвергающихся отбору. Из этого был сделан парадоксальный вывод о невозможности быстрых эволюционных преобразований, основанных на одновременном замещении многих независимых аллелей — которые, однако, реально происходили в филогенезе многих групп организмов. Этот парадокс решается, если учесть, что отбор действует, как уже подчеркивалось, не на отдельные гены, а на фенотипы организмов, формирующиеся в онтогенезе на основе взаимодействий целостных генетических систем («супергенов») в целостном генотипе. В результате отбору подвергаются не отдельные гены, а полигенные комплексы (супергены), и эволюционные преобразования затрагивают сопряженные комплексы фенотипических признаков. Это и определяет возможность высокой скорости эволюции без гибельных последствий для вида в результате слишком высокой платы за отбор (подробнее см. кн.: Грант В. Эволюция организмов. — М, 1980).
Полиморфизм вида Существующие в данный исторический момент условия среды могут благоприятствовать либо какому-то одному фенотипу, либо сразу нескольким разным фенотипам. В первом случае в природных популяциях преобладает один доминантный фенотип, называемый «диким типом», который скрывает в гетерозиготном состоянии многочисленные рецессивные аллели (таков, например, хорошо известный фенотип окраски домовой мыши). Во втором же случае в одной популяции постоянно встречаются несколько устойчивых, отчетливо различающихся друг от друга форм. Такое состояние называется полиморфизмом популяции (вида).
Очевидно, полиморфизм позволяет популяции лучше использовать разнообразие условий, существующих в пределах ее ареала аналогично тому, как дифференциация аллопатрических популяций позволяет виду лучше использовать разнообразие условий в пределах видового ареала. Полиморфизм повышает одновременно и приспособленность, и приспособляемость популяции, поскольку он всегда основывается на генетической гетерогенности, на существовании в популяции нескольких устойчивых генных комплексов.
Полиморфизм может и не иметь отчетливого приспособительного характера, основываясь на адаптивной равноценности речных вариантов (см. в гл. 1 о полиморфизме белковых комплексов). Но здесь нас интересует в первую очередь адаптивный, сбалансированный отбором полиморфизм. Такой полиморфизм в популяции возникает в следующих основных случаях: 1) отбор действует против фенотипа, обусловленного гомозиготным состоянием определенного аллеля, но благоприятствует гетерозиготному фенотипу, обладающему повышенной жизнеспособностью («сверхдоминирование» — гетерозиготы превосходят по жизнеспособности оба вида гомозигот); 2) на различных стадиях онтогенеза отбор благоприятствует фенотипическим особенностям, обусловленным разными аллелями; 3) отбор благоприятствует сохранению данного аллеля у особей одного пола, но действует против него у особей другого пола; 4) в разные сезоны года более приспособленными оказываются фенотипы, обусловленные разными аллелями; 5) фенотипы, обусловленные разными аллелями, используют различные условия в пределах одного ареала; 6) отбор благоприятствует сохранению данного фенотипа, когда этот последний редок, и направлен против него, когда он встречается часто.
Явления возрастного полиморфизма и полового диморфизма (вторая и третья формы полиморфизма в этом перечне) хорошо известны. Остальные варианты рассмотрим подробнее. Интересным примером «сверхдоминирования» является полиморфизм некоторых восточноафриканских популяций человека по типам гемоглобина. В этих популяциях у взрослых людей кроме обычного типа гемоглобина, А наблюдается также особая форма гемоглобина S, которая отличается от типа, А тем, что в одной из его молекулярных цепей место глютаминовой аминокислоты занимает валин. Нормальный аллель НbА встречается с частотой около 80%, мутантный аллель Hbs — около 20%. У гетерозигот НbА Hbs имеются обе формы гемоглобина, а у гомозигот HbsHbs почти весь гемоглобин крови представлен формой S. В этом последнем случае у людей развивается тяжелое заболевание — серповидно-клеточная анемия (эритроциты сморщиваются, принимая форму серпа), часто заканчивающаяся смертью. При таком исходе следовало бы ожидать гораздо меньшей частоты встречаемости мутантного аллеля, чем реально наблюдаемые 20%. Оказалось, что полиморфизм популяций по указанным аллелям связан с повышенной устойчивостью гетерозигот HbAHbs к малярии: количество паразитов в крови гетерозиготных индивидов много ниже, и продолжительность болезни короче, чем у гомозигот НbAНbА. Поскольку малярия является опасным заболеванием, вызывающим значительную смертность среди детей в Восточной Африке, очевидно, что распространение мутантного аллеля Hbs в человеческих популяциях в определенной концентрации будет выгодно, несмотря на опасность смертности гомозигот HbsHbs от анемии. В результате аллель Hbs имеет повышенную концентрацию именно в тех районах, где наиболее распространена малярия.
Наземные улитки Cepaea nemoralis в зоне смешанных лесов Умеренного пояса характеризуются сбалансированным полиморфизмом окраски раковины: выделяют бурую, розовую и желтую формы. Ранней весной наименее заметными на преобладающем фоне окружающей среды оказываются раковины бурой и розовой окраски, летом — желтые раковины. В то же время желтая форма менее заметна среди луговой растительности, тогда как бурая и розовая — в лесу. Очевидно, полиморфизм окраски у улиток соответствует 4-й и 5-й формам полиморфизма в приведенном выше перечне.
Наконец, последняя из указанных форм полиморфизма наблюдается, например, при мимикрии (подражательном сходстве) какого-либо вида с несъедобным для хищников видом-моделью. Такое сходство благоприятно для вида-подражателя, только если численность мимикрирующего фенотипа в его популяциях будет существенно ниже численности несъедобного вида-модели (в противном случае хищник не сможет эффективно «обучаться» не трогать особей с данным фенотипом). В ситуации, представленной на рис. 5, среди самок бабочки Papilio dardanus, разные цветовые устойчивые формы (морфы) имитируют окраску разных несъедобных бабочек-данаид, тогда как самцы P. dardanus вообще не мимикрируют.
Симпатрические внутривидовые группировки Полиморфизм популяций является основой для возникновения в некоторых случаях устойчивых и обособленных внутривидовых группировок особей («биологических популяций»), обитающих в одном и том же районе, т. е. обладающих симпатрическим распространением, в отличие от аллопатрических территориальных популяций. К ним относятся, в частности, возрастные и половые группы некоторых видов. Биологические популяции обособляются друг от друга, обитая в пределах одного ареала, либо в результате специфических взаимоотношений с внешней средой, либо благодаря особенностям внутривидовой организации. Примеры подобной внутривидовой дифференциации представляют обособленные друг от друга биологически личиночные и взрослые популяции земноводных или насекомых с полным превращением, или же такие связанные с особенностями размножения группировки, как «гаремы» и стада самцов-холостяков, характерные для целого ряда видов млекопитающих (кашалоты, олени, зебры, куланы и др.). раса генетический эволюция популяция Особой формой биологических популяций являются так называемые биологические расы. Этим термином обозначают такие устойчивые группировки особей одного вида, которые имеют симпатрическое распространение, но различаются друг от друга определенными экологическими особенностями (например, характером питания, сроками и местами размножения и т. п.). Благодаря этим различиям разные биологические расы одного вида держатся раздельно друг от друга, хотя и имеют общий ареал. В результате обмен генетической информацией между разными биологическими расами одного вида может быть снижен, как и между разными территориальными (аллопатрическими) популяциями.
Примером подобных биологических рас могут служить так называемые сезонные («яровые» и «озимые») расы ряда видов лососевых и осетровых рыб, описанные Л. С. Бергом. Речь идет о проходных рыбах, которые обитают в море, но для икрометания заходят в реки. Сезонные расы описаны Бергом у кеты (Oncorhyn-chus keta), чавычи (О. tschawytscha), нерки (О. nerka), семги (Salmo salar), каспийского лосося (S.trutta caspius), севрюги (Acipenser stellatus) и других видов. Рыбы «озимой» расы входят в реки осенью, зимой держатся в их низовьях, а рано весной поднимаются вверх по течению для нереста. Рыбы «яровой» расы проводят зиму в море и входят в реки позднее — когда нерест рыб «озимой» расы уже заканчивается. Благодаря этому обе расы одного вида, обитая в одном географическом районе и выметывая икру в одних и тех же реках, в значительной степени изолированы друг от друга во время сезона размножения и обмен генетической информацией между ними затруднен.
Другой пример представляют биологические расы обыкновенной кукушки (Cuculus canorus), описанные А. С. Мальчевским. Как известно, кукушка откладывает яйца в гнезда различных видов воробьиных птиц (это явление иногда называют «гнездовым паразитизмом»). Представители разных биологических рас обыкновенной кукушки проявляют предпочтение к «своему» виду воробьиных, в гнезда которого они откладывают яйца. При этом окраска яиц у кукушек разных рас соответствует окраске яиц того вида птиц, на котором «паразитирует» данная раса кукушек. Следовательно, в этом случае разные биологические расы проявляют уже Достаточно отчетливые наследственные различия морфологического признака (окраска яиц).
Очевидно, биологические внутривидовые группировки, так же как и территориальные популяции, позволяют виду более полно и совершенно использовать ресурсы в пределах занимаемого им ареала.
Подводя итоги, мы можем с полным основанием констатировать, что биологический вид представляет собой сложную систему, обладающую высокой устойчивостью к изменениям внешней среды и приспособленную к оптимальному использованию существующего разнообразия условий. Высокая степень приспособленности и приспособляемости вида обеспечивается несколькими путями, включающими мутационную, комбинативную и модификационную изменчивость и формирующиеся на базе комбинативной изменчивости политипичность и полиморфизм. Вид представляет собой систему с нежесткими связями взаимозаменяемых компонентов (так сказать, с «параллельным включением» звеньев), лишенную какого-либо интегрирующего, контролирующего всю систему центра. Эта сложная динамическая система возникает, поддерживается и совершенствуется под контролем естественного отбора.
Майкл Бамшед Стив Олсон Существуют ли расы?
Находясь на многолюдной улице мегаполиса, приглядитесь к беспрерывному потоку несущихся мимо людей, и вы увидите всё многообразие внешних признаков современного человечества. Перед глазами промелькнут лица с молочно-белой, жёлтой и тёмно-шоколадной кожей, шевелюры из прямых, волнистых или курчавых волос. Подобные морфологические признаки (наряду с географической областью происхождения и культурными традициями людей) мы чаще всего используем для подразделения всего человеческого рода на группы, именуемые расами. Но насколько оправданно понятие «раса» с биологической точки зрения? Несут ли биологические признаки, такие как цвет глаз, волос и кожи, достоверную информацию о генетической конституции человека? К тому же критерии определения расовой принадлежности в разных регионах планеты не одинаковы. Так, человека, которого в США назовут чёрным, в Бразилии сочтут за белого, а в Южной Африке — за цветного.
И тем не менее расхожие представления о расах порой оказываются эффективными при разграничении людей на группы в соответствии с их генетической предрасположенностью к определённым болезням. Серповидно-клеточная анемия, к примеру, наиболее широко распространена среди жителей Африки и Средиземноморья, а кистозный фиброз (муковисцидоз) — среди коренных европейцев. Результаты исследований указывают и на то, что у афро-американцев чувствительность к ряду лекарств, предназначенных для лечения сердечно-сосудистых заболеваний, намного меньше, чем у представителей других групп.
На протяжении последних лет учёные пристально изучали генетическую конституцию популяций людей в разных регионах планеты, пытаясь выявить связь между их происхождением и распространением различных заболеваний. Результаты исследований позволяют ответить на ряд вопросов, издавна вызывавших горячие споры в научном мире. Можно ли на основании генетической информации идентифицировать группы людей, имеющих общее происхождение, и определять принадлежность человека к той или иной группе? Будет ли такое распределение соответствовать разделению людей на группы на основании общепринятых представлений о расах? И, наконец, даёт ли разграничение (на основании традиционных принципов или генетического сходства) какую-либо полезную информацию о восприимчивости людей к той или иной болезни или их чувствительности к лекарственным препаратам?

На первый вопрос можно дать чёткий утвердительный ответ, на второй — чёткий отрицательный, а на третий — утвердительный ответ с некоторыми оговорками. Некоторые группы различаются генетически, но разграничение людей на группы зависит от того, какие именно гены изучаются. Проще говоря, благодаря природе генов, определяющих цвет кожи, вас могут отнести к одной группе, а по другим характеристикам — к другой. В многочисленных исследованиях было показано, что генетическое разнообразие людей на 90% обусловлено различиями, возникавшими внутри популяций, живших на каком-нибудь континенте, и лишь на 10% — различиями между континентальными популяциями. Иными словами, в среднем генетические различия в пределах какой-либо популяции людей выражены гораздо сильнее, чем различия между двумя популяциями. А это значит, что представители различных популяций отличаются друг от друга лишь чуть больше, чем представители одной и той же популяции.

Классификация людей.
100 тысяч лет назад люди современного анатомического типа вышли за пределы Африки и начали заселять другие части света. С тех пор численность нашего вида сильно увеличилась. Эти события сильно повлияли на структуру ДНК человека.
Для оценки степени родства между группами людей генетики используют мельчайшие различия (полиморфизмы) в структуре ДНК (например, различия в последовательности пар нуклеотидов). Как правило, полиморфизмы присутствуют не в генах, а потому прямого влияния на проявление тех или иных признаков не оказывают. Некоторые полиморфизмы, однако, затрагивают гены и поэтому могут усилить индивидуальные различия в проявлении внешних признаков, а также привести к развитию генетических заболеваний.
Расшифровав геном человека, учёные выявили миллионы полиморфизмов. Их распределение в популяциях людей отражает историю развития этих популяций и влияние на этот процесс факторов естественного отбора. Идеальным генетическим инструментом для разграничения групп людей мог бы быть полиморфизм, присутствующий в геноме всех представителей одной группы и отсутствующий у представителей всех других популяций. Однако такого полиморфизма, к сожалению, не существует: основные группы людей обособились сравнительно недавно и подверглись слишком сильному смешению.
Но для грубого распределения по группам можно воспользоваться полиморфизмами, возникающими с неодинаковой частотой в разных регионах планеты. Одно из семейств таких полиморфизмов составляют полиморфизмы Alu — короткие участки ДНК с одинаковой последовательностью нуклеотидов. Время от времени Alu реплицируются, и образующиеся копии непредсказуемым образом внедряются в новые сайты той же или другой хромосомы (присутствие Alu на новом месте, как правило, не отражается на функционировании соседних генов). Коль скоро произошла такая вставка Alu-элемента, он может оставаться на новом месте неопределённо долгое время, передаваясь из поколения в поколение. А значит, если два человека — носители одинаковой Alu-последовательности в одном и том же месте генома, то они должны иметь общего предка, наделившего их специфическими сегментами ДНК.
Обзор: Генетика рас.
Цвет кожи, текстура волос и прочие внешние признаки, на которых основаны традиционные представления о расах, определяются некоторой совокупностью генов. Но представители одной и той же расы могут значительно отличаться друг от друга другими генами. Соответственно, у представителей разных рас может быть большее генетическое сходство, чем у представителей одной и той же расы.
Исследователи научились использовать генетические данные для разграничения крупных популяций людей в соответствии с географией их происхождения. Этот подход, однако, не позволяет идентифицировать популяции, подвергавшиеся в прошлом интенсивному смешению с другими группами людей.
Споры по поводу значения расовых генетических различий для медицинской практики не утихают и сегодня.
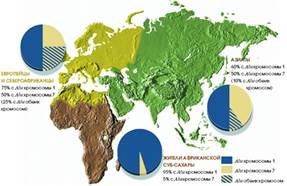
В исследовании, проведённом Майклом Бамшедом совместно с учёными из Университета штата Юта, Линном Джордем (Lynn B. Jorde), Стивеном Вудингом (Stephen Wooding) и Скоттом Уоткинсом (W. Scott Watkins), а также Марком Батцером (Mark A. Batzer) из Университета штата Луизиана, было изучено 100 различных полиморфизмов Alu у 565 человек, родившихся в Азии, Европе и регионах Африки, расположенных южнее Сахары. В первую очередь учёные определяли, является ли каждый из 565 обследованных человек носителем всех 100 Alu. Затем удаляли с полученных образцов биологического материала идентификационные этикетки (со сведениями о месте рождения и принадлежности к той или иной этнической группе) и «сортировали» людей по группам исключительно на основании генетической информации.
Проведённый анализ позволил выделить четыре группы людей. Когда биологические образцы вновь были снабжены своими опознавательными знаками, оказалось, что две группы целиком состояли из уроженцев Африки, причём одну из них почти полностью составляли пигмеи мбути. Каждая из двух других групп включала либо только европейцев, либо только уроженцев Восточной Азии. Как показали результаты исследования, чтобы установить с точностью до 90%, на каком континенте родился человек, достаточно изучить 60 полиморфизмов Alu. Анализ 100 полиморфизмов позволяет определить место происхождения человека с точностью до 100%.
Результаты исследований (см., например, статью Л. Животовского и Э. Хуснутдиновой «Генетическая история человечества» в № 7 журнала «В мире науки» за 2003 г.) показывают, что генетический анализ действительно позволяет разделять группы людей в соответствии с их географическим происхождением. Этот подход, однако, требует большой осторожности. Наиболее легко разграничиваются группы, сильно разобщённые географически и обнаруживающие наиболее выраженные генетические различия. Когда Бамшед воспользовался 100 полиморфизмами Alu, чтобы квалифицировать в качестве отдельной группировки выборку жителей Южной Индии, оказалось, что у индийцев гораздо большее генетическое сходство либо с европейцами, либо с азиатами. Другими словами, поскольку Индия испокон веков подвергалась сильному генетическому влиянию со стороны европейцев и жителей Восточной Азии, население этого субконтинента невозможно отнести к самостоятельному кластеру. Поэтому мы пришли к выводу, что для разграничения группировок, подвергавшихся в ходе исторического развития интенсивному смешению с другими многочисленными популяциями людей, необходим анализ многих сотен или даже тысяч полиморфизмов.
Расы и гены Если генетические данные позволяют подразделять людей на группы, возникает закономерный вопрос: отвечают ли общепринятые представления о расах определяющим их генетическим различиям? В некоторых случаях такое соответствие действительно существует, но так бывает далеко не всегда. Так люди, как правило, разделяют своих собратьев на расы на основании цвета кожи и черт лица, т. е. признаков, на формирование которых влияет естественный отбор. Но группы людей с одинаковыми физическими характеристиками, возникшими в результате естественного отбора, могут значительно различаться в генетическом отношении. Жители африканской суб-Сахары и австралийские аборигены могут иметь одинаковую пигментацию кожи (адаптация к сильному солнечному излучению), но отличаться генетически.
И наоборот: две генетически сходные группы могут подвергнуться воздействию неодинаковых факторов естественного отбора. Это может усилить некоторые различия между группами, и по внешнему виду их представители будут различаться гораздо сильнее, чем по генетической сути. Поскольку на формирование таких признаков, как, например, цвет кожи, влияет естественный отбор, они далеко не всегда отражают те популяционные процессы, которые были ответственны за распределение нейтральных полиморфизмов (типа Alu-последовательностей) или тандемных повторов. А следовательно, внешние признаки или полиморфизмы, подверженные действию естественного отбора, — не слишком надёжные критерии для распределения человека в ту или иную этническую группу, и заставляют иногда предполагать генетическое родство там, где его нет.
О том, насколько трудно бывает определить групповую принадлежность людей, свидетельствует и изучение различных популяций населения США. Большинство тех, кто считает себя афро-американцами, имеют предков, сравнительно недавно покинувших пределы Западной Африки, а по частоте полиморфизмов западные африканцы обычно значительно отличаются от европейцев, азиатов и коренных индейцев. Но доля западноафриканских полиморфизмов у афро-американцев сильно варьирует, т. к. на протяжении нескольких веков они подвергались интенсивному смешению с выходцами из различных областей Африки и иных частей света.
Генетическое разнообразие человечества Исследования, проведённые Марком Шрайвером (Mark D. Shriver) и Риком Киттлсом (Rick A. Kittles) из Университета Хауарда, позволили определить набор полиморфизмов, с помощью которого учёные оценивали у отдельных людей долю генов, происходящих из разных регионов планеты. Было установлено, что вклад западных африканцев в геном афро-американцев составляет 80%, но у разных жителей страны он варьирует от 20 до 100%. Эффект смешения групп проявляется и у многих людей, убеждённых в том, что их предки были исключительно европейцами. По данным Шрайвера, 30% американцев, считающих себя белыми, в действительности имеют менее 90% генов европейского происхождения. Таким образом, сведения, которые даёт большинство американцев о своём происхождении, далеко не всегда соответствуют их генетическому профилю. Как, впрочем, и расовая принадлежность человека, установленная на основании традиционных представлений о расах, далеко не всегда отражает его генетическую конституцию.
Гены, расы и болезни Понимание связи между расами и генетическими различиями имеет важное практическое значение. Некоторые полиморфизмы, частота которых в разных этнических группах различается, напрямую связаны со здоровьем людей. Так, мутации, ответственные за серповидно-клеточную анемию и некоторые формы кистозного фиброза (муковисцидоза), возникают в результате генетических изменений, особенно часто развивающихся у жителей Африки и Европы. Соответственно, они призваны защищать людей от распространённых в этих регионах болезней. У людей, перенявших только одну копию серповидно-клеточного полиморфизма, обнаруживается устойчивость к малярии, а люди с одной копией муковисцидозного полиморфизма менее предрасположены к обезвоживанию организма вследствие холеры. Сами же эти генетические заболевания возникают только у тех, кто унаследовал от родителей по две копии защитных мутаций.
Генетика для демографов Генетическими различиями можно объяснить и неодинаковую устойчивость людей к вирусу СПИДа. У некоторых счастливчиков имеются выпадения небольших участков ДНК в обеих копиях гена, кодирующего образование особых хемокиновых рецепторов (ССR5) на поверхности клеток. Соответственно, у них ССR5 на поверхности клеток отсутствуют. Поскольку большинство разновидностей вируса, вызывающего СПИД, могут проникать в клетки только после предварительного связывания с рецепторами ССR5, к ВИЧ-1-инфекции такие люди невосприимчивы. Полиморфизм в гене ССR5 обнаруживается почти исключительно у жителей Северо-Восточной Европы.
Некоторые полиморфизмы в гене ССR5 не препятствуют проникновению ВИЧ-1-инфекции, но влияют на сроки появления симптомов СПИДа и смерти больного. Одни из этих полиморфизмов порождают сходные эффекты во всех популяциях, где обнаруживается их присутствие, другие изменяют скорость развития инфекции лишь в некоторых группах. Так, например, один из подобных полиморфизмов замедляет прогрессирование болезни у евро-американцев, но ускоряет её развитие у афро-американцев. Сегодня учёные внимательно изучают подобные популяционные воздействия полиморфизмов в надежде применить полученные знания на практике — если, конечно, они научатся точно определять групповую принадлежность людей.
Знание «географических корней» имеет большое значение и для лечения заболеваний, широко распространённых в определённых популяциях людей. Гипертензия, диабет и многие другие болезни возникают в результате кумулятивного действия полиморфизмов в нескольких генах, по отдельности вызывающих лишь слабые патогенные эффекты. Кроме того, полиморфизмы, провоцирующие определённые патологические эффекты в одной группе людей, могут порождать совершенно иные эффекты у представителей другой популяции. Это затрудняет использование выявленных полиморфизмов в терапии генетических болезней. А до тех пор, пока учёные не оценят вклад генетических и средовых факторов в развитие подобных заболеваний, врачи, пытающиеся отыскать наилучший подход к их лечению, могут руководствоваться сведениями о «географических предках» своих пациентов.
Расы и медицина Однако в последние годы вопрос о связи между групповой принадлежностью людей и их здоровьем вызывает горячие споры среди учёных. В январе 2003 г. FDA США одобрило сбор данных о расовой и этнической принадлежности больных при проведении любых клинических испытаний. Но ряд исследователей настаивают на отмене рекомендаций FDA, а коллег, проводящих клинические испытания, они призывают сосредоточиться на сборе данных о геноме больных. Они считают, что различия между группировками людей настолько малы, а отрицательный исторический опыт, связанный с расовой дискриминацией, настолько велик, что при проведении генетических и медицинских исследований не должно придаваться серьезного значения групповой принадлежности больных. По мнению же других специалистов, оценить вклад генетических и средовых различий в развитие болезней невозможно без знания групповой принадлежности людей (включая и обычное определение рас на основе традиционных расовых характеристик, как, например, цвет кожи).
Кое-что о фотопортретах Серия фотопортретов на странице была создана нью-йоркской художницей Нэнси Берсон (Nancy Burson) с помощью изобретённой ею «Машины для моделирования человеческих рас». Это устройство изучает фотографию конкретного человека (в данном случае — белой женщины) и, изменяя его черты в соответствии с традиционными представлениями о расовых признаках, показывает, как он мог бы выглядеть, если бы относился к другой расе. «Машина» была одним из самых популярных экспонатов на лондонской выставке, посвящённой началу третьего тысячелетия, привлекавших к себе толпы посетителей. Более подробную информацию о художнице и её работах можно найти в сети Интернет на сайте www.nancyburson.com.
Значение расовой принадлежности для медицинских исследований обсуждалось и в мартовском выпуске журнала New England Journal of Medicine. Авторы одной из статей — Джей Кауфман (Jay S. Kaufman) из Университета Северной Каролины в Чапел-Хилл и Рик Уорд (Ryk Ward) из Оксфордского университета — уверены, что расовая принадлежность не является тем адекватным критерием, которым могли бы руководствоваться врачи при подборе лекарств для своих пациентов. Учёные ссылаются на два факта, выявленные при наблюдении за группами больных разной расовой принадлежности: 1. комбинация некоторых сосудорасширяющих препаратов оказалась гораздо более эффективной для лечения сердечной недостаточности у больных африканского происхождения, и 2. ингибиторы ангиотензин-превращающего фермента (АПФ) не вызывали у этих пациентов каких-либо выраженных терапевтических эффектов. А в другой статье группа исследователей, возглавляемая Нилом Ришем (Neil Risch) из Стэнфордского университета, отстаивает противоположную точку зрения: поскольку расовые или этнические группы людей скорее всего различаются генетически, расовые различия могут иметь большое значение при выборе терапевтического подхода. Авторы ссылаются на результаты исследования, показавшего, что частота осложнений при диабете II типа наиболее тесно коррелирует с расовой принадлежностью больных (даже если учитывается их уровень образования и доходов).
Во многих биомедицинских исследованиях принадлежность больных к той или иной группе определялась произвольно — на основании традиционных представлений о расах. Дискуссия о значении групповой принадлежности для клинической практики — наглядный пример того, насколько социальные и политические настроения в обществе влияют на формирование представлений о расах.
С незапамятных времён лучшие умы человечества задавались вопросом, откуда взялись различные группы людей и в каком родстве они состоят друг с другом. Учёные и философы строили догадки о том, почему их представители столь сильно различаются внешне и ограничиваются ли биологические различия между людьми только цветом кожи, разрезом глаз и другими внешними признаками. Новые генетические данные и последние методы анализа наконец-то приблизили нас к решению многих из этих загадок. Они помогут нам лучше понять собственную биологическую природу и осознать неразрывность родственных уз, объединяющих всех людей в единую генетическую семью.
Об авторах:
Майкл Бамшед (Michael J. Bamshad) — генетик из Медицинской школы при Университете штата Юта — исследует генетику популяций в надежде лучше понять историю человечества и происхождение различных заболеваний.
Стив Олсон (Steve E. Olson) — писатель, автор нашумевшей книги Mapping Human History: Genes, Race, and our Common Origins («Картирование истории человечества: гены, расы и наши общие корни»), выдвинутой в 2002 г. на соискание ежегодной Национальной книжной премии.