Царство Грибы.
Царство Грибы

Бурая листовая ржавчина пшеницы (P. triticina). Начальный этап развития гриб проходит на видах рода василистник (Thalictrum), затем поражается пшеница. Этот гриб редко проходит полный цикл развития. Зимует он в виде урединиоспор или дикариотичного мицелия на озимой пшенице. Таким образом, он способен не один год существовать в дикарио фазе. Такую ситуацию можно считать результатом своеобразной… Читать ещё >
Царство Грибы. Царство Грибы (реферат, курсовая, диплом, контрольная)
Настоящие грибы — это организмы самого разнообразного строения. Чаще всего вегетативное тело — мицелий, обычно септированный, многоклеточный, однако самые примитивные формы одноклеточные или имеют плазмодий (ризоминелий). Клеточные стенки, как правило, содержат хитин, маннан и глюканы, другие вещества, в частности целлюлоза, встречаются лишь изредка. Митохондрии имеют пластинчатые кристы. В отличие от грибоподобных организмов (микоидов) жгутики у спор (зооспор) известны только у самых примитивных представителей. Обычно это один гладкий жгутик, направленный назад по ходу движения зооспоры или гаметы. Для многих настоящих грибов характерно наличие в цикле развития дикариофазы, завершающейся образованием мейоспорангиев, возникающая при этом диплофаза очень короткая.
Экология грибов чрезвычайно разнообразна: здесь есть и сапротрофы, и паразиты, и симбионты. Роль этих организмов в природе колоссальна, прежде всего как редуцентов. Мертвый растительный материал разлагается почти исключительно настоящими грибами.
Царство настоящие грибы обычно подразделяется на несколько отделов.
Низшие грибы:
Хитридиомицеты (Chytridiomycota). Мицелий развит слабо или вегетативное тело представляет собой одиночную, иногда лишенную стенки клетку (плазмодий). Имеются формы с ризомицелием. Бесполое размножение с помощью зооспор с одним бичевидным («гладким») жгутиком. Половой процесс — гаметогамия разных типов или хологамия. В клеточных стенках содержатся хитин и глюканы. Только у хитридиомицетов имеются жгутиковые стадии.
Зигомицеты (Zygomycota). Мицелий хорошо развит, у большинства представителей отдела он неклеточный. Дрожжевидные формы известны, но встречаются редко и лишь в определенных условиях. Бесполое размножение преимущественно спорангиоспорами, реже односпоровые спорангии отрываются целиком и таким образом функционируют как конидии. Половой процесс — зигогамия. Клеточные стенки мицелия содержат хитин и хитозан.
Высшие грибы:
Аскомицеты (Ascomycota). Мицелий хорошо развит, септированный. Встречаются также дрожжевидные формы. Бесполое размножение при помощи конидий. Половой процесс — чаще всего гаметангиогамия. Споры полового размножения образуются эндогенно — в сумке. Клеточные стенки содержат хитин и глюканы, у дрожжей — глюканы и маннаны.
Базидиомицеты (Basidiomycota). Мицелий хорошо развит, септированный. Известны дрожжевидные формы. Бесполое размножение при помощи конидий. При половом процессе никаких специальных половых органов не образуется. Споры полового размножения образуются экзогенно — на базидии. Клеточные стенки содержат хитин и глюканы.
Как отдельная группа выделяются несовершенные грибы, или дейтеромицеты, представляющие собой совокупность конидиальных стадий (анаморф) аскомицетов (значительно реже базидиомицетов), у которых половое спороношение неизвестно, встречается редко или в другое время (например, после зимовки).
Класс базидиомицеты — Basidiomycota.
Общая характеристика:
Базидиомицеты — высшие грибы с многоклеточным мицелием. К ним относятся около 30 тыс. видов (и микроскопические грибы, и грибы с крупными плодовыми телами). Среди этих грибов есть паразиты растений (например, широко распространенные и очень опасные для сельскохозяйственных растений головневые и ржавчинные грибы), многочисленные почвенные сапрофиты — хорошо всем известные шляпочные грибы (например, шампиньоны, навозники). К базидиомицетам относятся образующие шляпочные грибы, которые успешно развиваются только в тесном контакте с корнями древесных растений (например, белый, подберезовик, подосиновик и многие другие лесные грибы).
Есть среди базидиальных грибов и сапрофиты на древесине — это многочисленные трутовики — активные разрушители древесины и валежника.
Половое спороношение у них — базидио споры, т. е., экзогенные споры на особых выростах — базидиях. Такая базидия закладывается из двух ядерных клеток. Половых органов нет. Половой процесс осуществляется путем слияния двух вегетативных клеток гаплоидного мицелия, вырастающего из базидио споры и состоящего из одноядерных клеток. У гомоталличных видов могут сливаться гифы одного и того же мицелия. У гетероталличных, к которым относится большинство базидиальных грибов, сливаются клетки гиф, берущих начало от спор противоположных половых знаков: + и -. При этом происходит слияние цитоплазмы, а ядра объединяются в пары — дикарионы, которые затем делятся синхронно. Такой дикариофитный мицелий (с двух ядерными клетками), пронизывая субстрат (почву, древесину, стебли и листья растений-хозяев), может существовать длительное время. У некоторых базидиальных грибов, например у трутовиков, растущих на деревьях, или у лесных шляпочных грибов, мицелий многолетний.
На концах дикариофитных гиф из двухъядерных клеток образуются базидии. На базидии развиваются 2−4 базидио споры, сидящие на маленьких шипообразных выростах базидии — стеригмах. Дикариофитный мицелий у большинства видов базидиальных грибов характеризуется наличием пряжек, особых клеточек у поперечной перегородки клеток мицелия. Базидии с базидио спорами могут возникать прямо на мицелии. Но у большинства базидиомицетов они образуются на плодовых телах или внутри них.
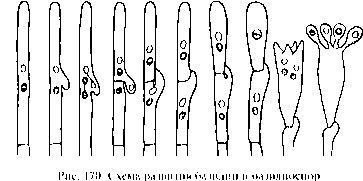
Рассеивание базидио спор происходит путем их активного отбрасывания. В основе этого лежит повышение в самой базидии внутриклеточного давления в результате гидролиза гликогена. Процесс гидролиза сопровождается притоком в базидию воды, в результате чего и повышается в ней давление. Оно передается к базидио споре через узкий канал стеригмы, что значительно его ослабляет. В результате зрелая базидио спора получает незначительный толчок при отбрасывании и отлетает всего на несколько десятых миллиметра. В дальнейшем она подхватывается токами воздуха. При массовом отбрасывании базидио спор у шляпочных грибов и трутовиков часто часть этих спор заносится на поверхность шляпки, и тогда можно наблюдать, как какая-либо часть шляпки, например у красной сыроежки, покрыта белым бархатистым налетом ее базидио спор, а сероватая, например, шляпка шампиньона покрыта таким же бархатистым налетом шоколадно-коричневых базидио спор. В закрытых плодовых телах (у гастеромицетов) базидио споры не отбрасываются так активно. Они освобождаются в результате разрушения базидий и общей оболочки плодового тела, а затем разносятся токами воздуха.
Плодовые тела сложены из дикариофитного мицелия. Следовательно, в цикле развития базидиальных грибов преобладает дикариофитный мицелий.
Гаплоидная фаза короткая: базидио споры и мицелий, выросший из нее и существующий небольшой период. Конидиальные спороношения (бесполое размножение) у базидиальных грибов встречаются редко. Плодовые тела базидиомицетов различны по форме и консистенции. Они могут быть паутинистыми, рыхлыми, плотно-войлочными, кожистыми, деревянистыми, мягко-мясистыми, могут иметь форму пленок, корочек, могут быть копытообразными или состоять из шляпки и ножки.
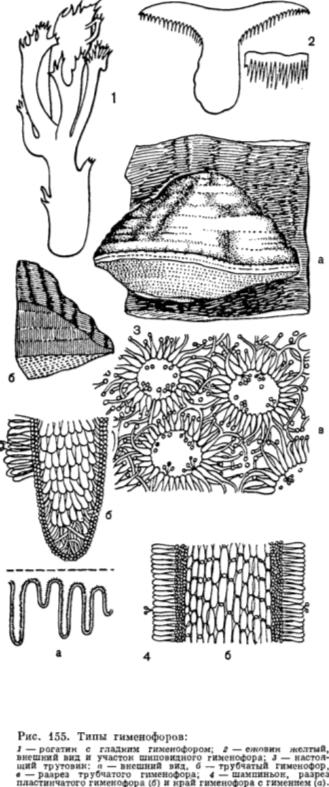
Спороносный слой плодового тела — гимений — располагается у более примитивных видов на верхней стороне плодовых тел, а у более высокоорганизованных — на нижней. Гимений базидиальных грибов состоит из базидий с базидио спорами и парафиз. У некоторых видов в гимении находятся цистиды — крупные клетки, возвышающиеся над гимениальным слоем. Они защищают гимениальный слой и особенно базидий от давления сверху. Форма цистид для многих видов постоянна и часто служит признаком для их определения.
Поверхность плодового тела, несущую гимении, называют гименофором. У низших представителей он гладкий, а у более высокоорганизованных имеет форму зубцов, трубочек, пластинок.
Классификация базидий:
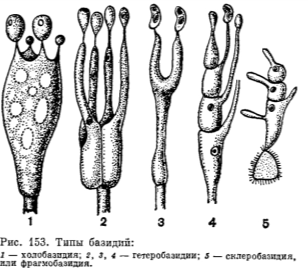
Холобазидия (голобазидия): не разделена перегородками.
Фрагмобазидия: разделена перегородками (септирована).
Гомобазидия (хомобазидия) и гетеробазидия. Гомобазидия — это разросшаяся материнская клетка. Если материнская клетка превращается как бы в первый этаж двухэтажной конструкции, а вторым этажом («надстройкой») служит специальный вырост (эпибазидия), на котором непосредственно формируются базидио споры, то это гетеробазидия.
Если гетеробазидия развивается из специальной покоящейся споры, закономерно входящей в цикл развития гриба (так называемой телио споры), то ее называют телиобазидией. Стихобазидия и хиастобазидия. Это различие обусловлено ориентацией оси веретена первого деления диплоидного ядра в формирующейся базидии. В стихобазидии веретено ориентировано параллельно ее оси, в хиастобазидии — перпендикулярно.
Классификация класса базидиомицетов.
Имеются различные варианты классификации, рассмотрена одна из них («компромиссный вариант»).
Выделяют 3 подкласса:
1. Урединиомицеты (Urediniomycetes), или телиомицеты (Teliomycetes). Разделенная перегородками базидия (фрагмобазидия) развивается из покоящейся споры (телио споры), возникающей как специальное образование (а не в результате обособления отдельных частей ранее существовавшего мицелия). Септы простые, утончающиеся к середине поры. Парентосомы (поровой колпачок с отверстиями) отсутствуют. Представители подкласса — в основном паразиты растений, реже насекомых, очень немногие — сапротрофы.
К ним относятся порядки:
- 1) Ржавчинные — Uredinales;
- 2) Септобазидиевые — Septobasidiales.
- 2. Устилагиномицеты (Ustilaginomycetes), или устомицеты (Ustomycetes). Базидия, разделенная перегородками или цельная, также развивается из покоящейся споры, но последняя возникает из ранее существовавшего мицелия. Септы простые, равномерной толщины или утолщенные к поре. Многослойных парентосом нет, но поры часто прикрыты колпачками или дисками обычно мембранного происхождения.
Порядки:
- — Головневые — Ustilaginales;
- — Экзобазидиевые — Exobasidiales.
- 3. Собственно базидиомицеты (Basidiomycetes). Основная часть подотдела. Базидия развивается от мицелия, чаще всего на специальных плодовых телах (базидиомах). В большинстве случаев она является цельной (голобазидией), не состоящей из гипои эпибазидии, т. е., гомобазидией. Септы долипоровые, парентосомы разной конфигурации.
Надпорядок I. Гетеробазидиомицеты — Heterobasidiomycetidae.
Порядки:
- 1) Тюланелловые — Tulasncllalcs;
- 2) Аурикуляриевые — Auriculariales;
- 3) Дакримицетовые — Dacrymycetalcs.
Надпорядок II. Дрожалковые, или Тремемовые — Tremellomycetidae.
Порядки:
- 1) Тремелловые — Tremellales;
- 2) Филобазилисвые — Filobasidiales.
Надпорядок III. Гомобазидиомицеты — Homobasidiomycetida.
Порядки:
- 1) Лисичковыс — Cantharcllales;
- 2) Пориевые — Poriales;
- 3) Гименохетовыс — Hymenochactales;
- 4) Ганодермовые — Ganodcrmatales;
- 5) Телефоровые — Thelcphorales;
- 6) Полипоровые — Polyporales;
- 7) Болетовые — Boletales;
- 8) Сыроежковые — Russulales;
- 9) Агариковые — Agaricalcs.
Гастероидные:
- 1) Дождевиковые — Lycoperdales;
- 2) Геастровые, или земляные звездочки, — Geastrales;
- 3) Склеродсрмовые, или ложнодождевиковые, — Sclerodermatales;
- 4) Тулостомовые — Tulostomatales;
- 5) Гнсздовковые — Nidulariales;
- 6) Веселковые — Phallales.
Вегетативное тело — развитый септированный мицелий. Встречаются дрожжевидные стадии или формы. Строение септ в мицелии разнообразно и существенно отличается от простых неслоистых септ аскомицетов.
Вегетативное размножение — фрагментацией мицелия или почкованием дрожжевых клеток. Бесполое (споровое) размножение — только конидиями, но встречается у базидиомицетов относительно редко, причем обычно наряду с половыми спороношениями. Только у представителей одного порядка ржавчинные (Uredinales) широко распространены специфические бесполые споры, имеющие специальные названия (далее подробно).
Половой процесс всегда происходит без образования специальных половых структур — соматогамия. Чаще всего это осуществляется путем контакта гаплоидных мицелиев или же оплодотворяющую функцию берут на себя спермации, образующиеся по типу конидий — сперматизация.
При половом процессе, происходит координация ядер в дикарионы с последующим их синхронным делением.
В результате полового процесса образуется базидия. Эта структура соответствует мейоспорангию, но мейоспоры, называемые здесь базидио спорами, образуются на ее поверхности, т. е. — экзогенно. При развитии базидии происходит кариогамия (слияние двух ядер дикариона), мейоз и переход гаплоидных ядер в базидио споры. Базидии могут быть цельными или разделены перегородками. У некоторых базидиомицетов базидии развиваются из специальных спор.
Половой процесс и образование базидии с базидио спорами у очень многих видов сопряжены с развитием плодовых тел самых разных размеров и облика.
В жизненном цикле явно преобладает дикарио фаза. Гаплофаза сильно подавлена, а дикариотичный мицелий может жить несколько столетий. В частности, дикариотичным мицелием сложены плодовые тела. У некоторых представителей отдела отмечена диплоидность.
Базидиомицеты широко распространены в природе во всех географических областях, на самых разных субстратах. Среди них очень много сапротрофов, паразитов растений и микоризообразователей. Такие базидиомицеты, как паразиты животных и человека, микобионты лишайников, а также обитатели водной среды, известны, но их очень мало. Очень многие базидиомицеты — съедобные грибы, широко используемые в пищу. Некоторые из них культивируются. Ряд видов используется в экспериментальных биологических исследованиях.
Далее будут рассмотрены (в соответствии с темой реферата) только порядки и виды паразитов культурных злаковых.
Базидиомицеты — паразиты культурных злаковых.
Порядок ржавчинные — Uredinales.
Все представители этого обширного (более 5 000 видов) порядка — облигатные биотрофные паразиты самых разных высших растений. Они вызывают характерные пятна или полосы разных тонов ржаво-бурого цвета, очень напоминающие поражение черного металла ржавчиной (откуда и название). Пятна или полосы могут быть довольно светлые, оранжевые (как свежая ржавчина), коричневые (как ржавчина более старая) и, наконец, почти черные (как очень старая ржавчина). Они представляют собой спороношения, которых у ржавчинных грибов может быть несколько и которые очень непохожи одно на другое. Это грибы, имеющие эндофитный, межклетный мицелий (местный или диффузный) с гаусториями, внедряющимися внутрь клеток хозяина. Мицелий содержит капли масла, окрашенные пигментом в оранжевый цвет. Такие же капли масла имеются в спорах. Именно они обусловливают «ржавые» цвета спороношений.
В цикле развития ржавчинных могут последовательно чередоваться внешне совершенно разные спороношения (имеющие наряду с разной морфологией и разную окраску). Следует отметить, что установление связи разных спороношений между собой было делом достаточно трудным, и далеко не во всех случаях эти связи установлены и до сих пор. У некоторых ржавчинных весь жизненный цикл проходит на одном и том же круге растений-хозяев, у других гаплоидный мицелий с соответствующими спороношениями развивается на одном круге растений-хозяев, а дикариогичный, также с соответствующими спороношениями, — на другом.
Некоторые типы спороношений могут отсутствовать. При этом у одних видов бывает как полный цикл (со всеми спороношениями), так и неполный (при отсутствии некоторых из них). У других видов полного цикла не бывает.
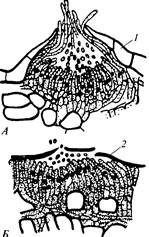
Базидии, разделенные поперечными перегородками (фрагмобазидии), образуются при прорастании покоящихся спор (телио спор) после периода покоя (в районах умеренною климата — после зимовки). Телио споры могут быть одноклеточными или же состоять из двух или большего числа клеток. В любом случае каждая клетка дикариотична. Перед прорастанием телио спор происходит сначала кариогамия — слияние двух ядер дикариона с образованием диплоидного ядра. Затем следует мейоз. Гаплоидные ядра по одному переходят в каждую клетку базидии, а из них — в формирующиеся базидио споры.
Базидио споры формируются на стеригмах, образуют капельки и за счет сил поверхностного натяжения могут отбрасываться на некоторое расстояние. Затем они распространяются воздушными течениями. Попав на соответствующее восприимчивое растение (обычно на лист), они прорастают, проникая в ткань. Начинается развитие гаплоидной стадии гриба. На гаплоидном мицелии, состоящем из одноядерных клеток, возникают пикнии — спороношения, напоминающие перитеции аскомицетов; отличаются они только тем, что в их полости расположены короткие спорогенпые клетки — фиалиды, образующие очень мелкие шаровидные споры — по типу образования фиалоконидии. Однако задача этих спороношений — произвести оплодотворение. Ржавчинные грибы, насколько известно, все гетероталличны, поэтому для дикариотизации необходима координация ядер разных половых знаков. Это может произойти при непосредственном контакте в пораженном органе мицелиев разных знаков, но существует и другой способ дикариотизации — перенос спермациев с мицелия одного знака на мицелий другого знака.
Пикнии закладываются как шаровидные клубки гиф под эпидермисом растения-хозяина, давят на него, прорывают и высовывают наружу пучок коротких гиф — перифиз. Между ними выступает наружу сахаристая пахучая жидкость (один вид ржавчинных грибов поэтому так и называется: пукциния пахучая — Puccinia suaveolens) с массой погруженных в нее спермациев. Эта жидкость привлекает насекомых, переносящих спермации с одного спермогония на другой, где они прилипают к перифизам и сливают с ними свое содержимое (если, конечно, тот спермогоний образован мицелием другого знака). После дикариотизации развиваются новые спороношения — эции. При поражении листа они обычно приурочены к его нижней стороне. В них образуются эциоспоры, которые развиваются всегда цепочками, образующими плотный палисадный слой, и содержат каждая по два несестринских ядра дикариона.
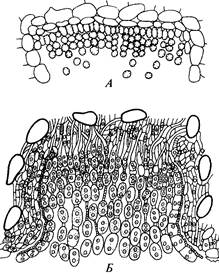
Каждая цепочка берет начало от одной клетки, называемой базальной.
Базальная клетка отбрасывает сначала одну дикариотичную клетку, потом другую, далее — третью и т. д. Это материнские клетки эциоспор, каждая из которых делится на две неравные по размеру клетки. Дальняя от базальной клетки, более крупная, формирует относительно толстую оболочку и превращается в эциоспору, ближняя, промежуточная (дизъюнктор), остается тонкостенной, впоследствии она разрушается и служит для разделения эциоспор. У многих ржавчинных грибов краевые базальные клетки, располагающиеся кольцом вокруг всего палисадного слоя цепочек, также отчленяют цепочки двуядерных клеток, которые срастаются между собой и формируют однослойную оболочку — перидий. Конечные эциоспоры срастаются между собой и с боковыми стенками, образуя крышу перидия. Нарастающие основанием базальные клетки и увеличивающиеся в размере эциоспоры разрывают эпидермис и крышу перидия, края которого отворачиваются в виде лопастей. Созревшие эциоспоры выпадают и распространяются токами воздуха.
Различают несколько типов эциев:
- 1) тип эцидиум (Aecidium) — спороношение шаровидной формы, масса спор окружена перидием, т. е., оболочкой из сросшихся недоразвитых эциоспор;
- 2) тип цеома (Caeoma) — перидия нет;
- 3) тип рестелин (Roestelia) — перидий долго продолжает расти своим основанием, так что получается нередко длинный рожок, высовывающийся из пораженной ткани;
- 4) тип перидермиум (Peridermium) — перидий имеется, но само спороношение закладывается в виде плоского, нарастающего краями сплетения гиф. На этом этапе некоторые виды вызывают различного рода видоизменения пораженных частей растений: опухоли, искривления и т. д., в том числе «ведьмины метлы», напоминающие подобные же образования при поражении растений тафриновыми грибами. У молочая при поражении грибом Uromycespisi форма листьев вместо обычной узколанцетной бывает почти округлой, как у брусники, такие растения даже описывали как самостоятельный вид.
Если ржавчинный гриб, то на этапе образования эциоспор происходит смена хозяев: эциоспоры не могут заразить ни одно растение из тех, на которых развивался гаплоидный мицелий со спермогонями и эциями. В этом случае эциоспоры должны попасть на растение из совершенно другого круга хозяев. На нем эциоспоры прорастают, проростковые трубки через устьица проникают в ткани, где формируется межклеточный дикариотичный мицелий с гаусториями. На мицелии развиваются летние спороношения гриба — урединии, представляющие собой скопления одноклеточных овальных урединиоспор, сравнительно тонкостенных, сидящих на ножке и содержащих по два ядра дикариона.
Закладываются эти спороношения, как и предыдущие, под эпидермисом листа. Развиваясь и увеличиваясь в размере, урединиоспоры давят на эпидермис, разрывают его и освобождаются. Они отрываются от ножек, рассеиваются в воздухе и могут снова заражать растения, где, как и при заражении эциоспорами, развивается дикариотичный мицелии, каких новых поколении за вегетационный период обычно бывает несколько. При наступлении неблагоприятных условий урединиоспоры могут превращаться в телиспоры.
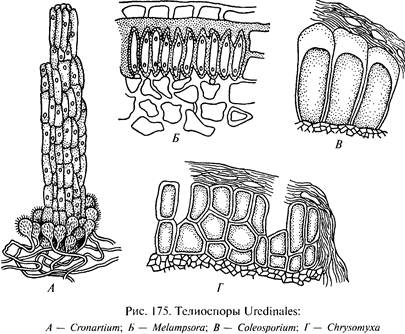
К концу периода вегетации на том же дикариотичном мицелии образуются новые спороношения — телии, представляющие собой скопления телио спор, или зимних спор. Оболочка у них толстая, почти черная. Предназначены телио споры для сохранения гриба в зимний период. Они могут быть одноклеточными, двух клеточными или состоять из большего числа клеток. У одних грибов телио споры на ножках, как и урединиоспоры, у других ножек нет, а споры соединяются в корочки, цепочки или колонки. После периода покоя телио споры прорастают в базидии.
Полный цикл ржавчинного гриба замыкается.
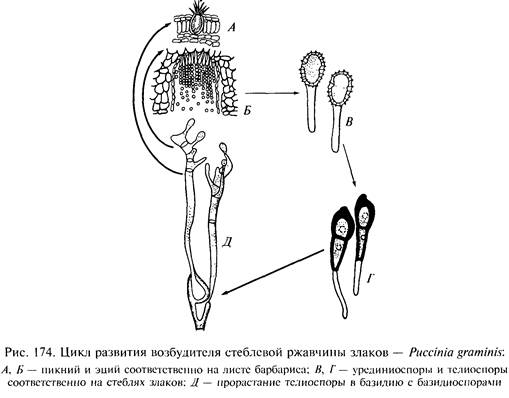
Таким образом, в полном цикле ржавчинных грибов последовательно сменяются пять типов спороношений:
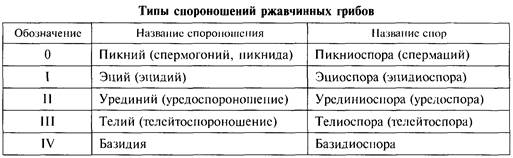
Представители — паразиты культурных злаковых:
I. Род Puccinia характеризуется также двух клеточными телио спорами, но ножки их не ослизняются, а студенистая масса не образуется Эции с перидием. Это самый большой по числу видов род в порядке ржавчинных грибов: он насчитывает свыше 2 000 видов.
1) Наиболее известен вид P. gramlnis — возбудитель стеблевой ржавчины злаков (ее называют также линейной или черной ржавчиной). Пикнии и эции этого гриба развиваются на представителях семейства барбарисовых (Berberidaceae) — видах рода барбарис (Berberis) и магония (Mahnia). Эциоспоры заражают самые различные злаки — как дикорастущие, так и хлебные, на которых развиваются урединии и телии. Именно на этом виде был впервые прослежен полный цикл развития разнохозяинных ржавчинных грибов.
Хотя P. graminis характеризуется полным циклом развития, прохождение всех стадий необязательно. Например, возможна перезимовка дикариотичнго мицелия в корневищах дикорастущих злаков с последующим переходом весной на культурные растения.
При исследовании P. graminis было открыто также существование специализированных форм и физиологических рас, впоследствии установленное у многих других фитопатогенных грибов. Было обнаружено, что вид P. graminis состоит из ряда специализированных форм (forma specialis), почти неотличимых по морфологии (размерам и форме спор), но способных заражать лишь определенные виды злаков. Так, пшеничная форма (P. graminis f. sp. tritici) паразитирует на пшенице, редко на ржи, ржаная (P. graminis f. sp, secalis) — на ржи, ячмене и некоторых видах пырея, овсяная (P. graminis f. sp. avenae) — на овсе и некоторых других представителях трибы Avenae, P. graminis f. sp. phleipratensis — на тимофеевке, лисохвосте и др.
Все эти формы в типичном случае имеют полный цикл развития, однако каждая из них обладает некоторыми экологическими особенностями, связанными с особенностями растений-хозяев. Так, P. graminis f. sp. secalis может зимовать в виде мицелия в корневищах некоторых видов пырея и с них переходить на рожь и ячмень, P. graminis f. sp. tritici более других связана с видами барбариса и часто заражает пшеницу, попадая сначала на дикорастущие злаки.
Вид P. graminis послужил хорошим объектом для изучения миграции фитопатогенных организмов на очень большие расстояния. Так, было показано, что в Западной Канаде источником раннего первичного заражения пшеницы служат эциоспоры и урединиоспоры с юга США и даже севера Мексики. Осенью происходит обратное явление. Урединиоспоры на юге США (например, в Техасе) часто гибнут во время жаркого лета, но молодая озимая пшеница может в начале осени заражаться урединиоспорами, принесенными за сотни километров из зараженных ржавчиной более северных районов.
- 2) Бурая листовая ржавчина пшеницы (P. triticina). Начальный этап развития гриб проходит на видах рода василистник (Thalictrum), затем поражается пшеница. Этот гриб редко проходит полный цикл развития. Зимует он в виде урединиоспор или дикариотичного мицелия на озимой пшенице. Таким образом, он способен не один год существовать в дикарио фазе. Такую ситуацию можно считать результатом своеобразной адаптивной эволюции: изменение биологии гриба в связи с особенностями развития его хозяина. Зимовка на озимых позволяет грибу почти постоянно находиться на живых растениях, а не на растительных остатках, где телио споры в период с осени до весны часто разрушаются микроорганизмами. Кроме того, виды василистника (растения, тяготеющие к более или менее сырым местам) не всегда имеются у гриба «под рукой». Прохождение время от времени полного цикла развития обеспечивает грибу генетическое разнообразие и в связи с этим дает возможность приспосабливаться к новым для него сортам.
- 3) Желтая ржавчина (P. striiformis) известена только в дикариофазе, зимует на озимой пшенице.
- 4) Корончатая ржавчина овса (Р. соronifera). Гаплофаза развивается на жестере слабительном (крушине слабительной — Rhamnus cathartica), дикарио фаза — на овсе и некоторых других представителях трибы Avenae. Этот гриб обычно проходит полный цикл, поэтому наибольший вред он приносит в районах произрастания жестера.
Порядок головневые — Ustilaginales. Все головневые грибы, как и ржавчинные, облигатные биотрофные паразиты многих цветковых растений. Они вызывают характерные поражения тех или иных частей растения (очень часто генеративных, но нередко поражаются листья и стебли, а иногда и корни). Пораженные части выглядят как бы обгорелыми, обугленными (отсюда название болезни — головня, соответственно и название грибов). Связано это с тем, что внутри тканей растения из вегетативного мицелия развиваются в массе очень темные споры. По месту в жизненном цикле это телио споры. Для них применяется специальное название — головневые споры, или усто споры. Они бывают одиночные, обособленные при созревании, или срастающиеся друг с другом в спорокучки. Такие спорокучки могут состоять только из собственно телио спор, но дополнительно могут иметь светлые стерильные клетки, неспособные к прорастанию. Особенно крупные спорокучки у головневых грибов, поражающих водные растения — видов родов Doassansia и Doassansiopsis. Эти спорокучки распространяются в воде (подобно плодам кокосовой пальмы), а имеющие газообразное содержимое стерильные клетки выполняют функцию понтонов.
Мицелий у головневых грибов дикариотичный, эндофитный, большей частью межклетный, диффузный или локальный. Он распространяется по межклетникам тканей зараженных растений, проникая в клетки при помощи гаусториев. Нахождение мицелия в тканях поначалу мало сказывается на внешнем облике растений. Иногда он может даже стимулировать их развитие, и только когда начинается формирование усто спор, появляются и внешние симптомы поражения. В некоторых случаях скопление усто спор на вегетативных органах становится заметным вскоре после заражения растений, например при заражении кукурузы пузырчатой головней.
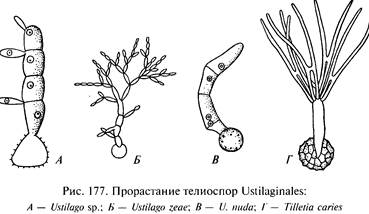
После освобождения из разрушенной ткани хозяина усто споры обычно способны к немедленному прорастанию, но могут годами сохранять всхожесть. При прорастании они развивают базидии без перегородок или с 1 — 3 перегородками. При образовании базидио спор гаплоидное ядро в каждой клетке разделенной базидии делится и одно получившееся новое ядро идет в базидио спору, а другое остается в базидии и может делиться снова, за счет чего могут образовываться новые базидио споры. Циклы развития головневых грибов, различаясь в деталях, имеют одну общую особенность: заражение происходит только в дикарио фазе.
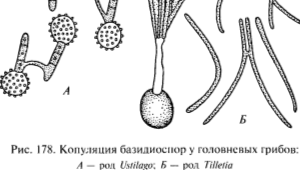
Чаще всего это происходит следующим образом. Усто споры, содержащие по 2 ядра дикариона, попадают в почву с зараженными спорами посевными зернами. Здесь они прорастают в базидию, в которую после мейоза поступают гаплоидные ядра, переходящие в базидио споры. На базидии развиваются 2 споры со знаком «+» и 2 — со знаком «-». Базидио споры непосредственно копулируют между собой с образованием копулятивных мостиков или сначала почкуются, а затем копуляция происходит уже между производными клетками.
После копуляции развивается дикариотичный мицелий. Он заражает растение, прорывая ткани молодых проростков до их выхода на поверхность почвы. Затем он проникает в конус нарастания и продвигается вверх, пронизывая узлы и междоузлия.
Когда растение-хозяин становится достаточно взрослым, мицелий в определенных местах распадается на отдельные клетки, одевающиеся темной оболочкой, усто споры. По этому типу развиваются: Tilletia caries, Т. secalis, Sphacelotheca panici-miliacei, Uwcystis occulta, Ustilago horde и многие другие головневые грибы.
У некоторых других головневых грибов цикл проходит несколько иначе.
Например, усто споры возбудителя пыльной головни пшеницы (Ustilago tritici) и ячменя (U. nuda) для дальнейшего развития должны попасть на рыльце цветка восприимчивого растения-хозяина, где они прорастают четырехклеточной базидией. Происходит мейоз, в результате которого возникает четыре гаплоидных ядра. Однако базидио споры не развиваются, а дикарион возникает путем перехода ядер из одной клетки базидии в другую или слияния соседних клеток. Если ядра разного знака находятся не в соседних клетках, могут образовываться специальные мостики. Мостики могут возникать также между клетками разных базидии. После образования дикариона из базидии вырастает дикариотичный мицелий, по пыльцевой трубке проникающий в завязь. При этом внешне семя выглядит совершенно нормально.
Попав в почву, такие семена без видимых отклонений прорастают. Молодое растение развивается, и в нем развивается дикариотичный мицелий гриба. Он проникает в закладывающийся колос, обильно там разрастается, разрушает формирующиеся колоски и затем распадается на усто споры. В вышедшем из листового влагалища колосе сохраняются только стержень и сильно разрушенные колоски. Все остальное превращается в темную пылящую массу усто спор, способных прорастать без периода покоя. Они переносятся ветром на завязи цветущих в это время колосьев. Цикл замыкается.
Специфику такого жизненного цикла необходимо учитывать при борьбе с соответствующими патогенами.
Обычно в качестве меры химической борьбы с головневыми грибами применяют протравливание, т. е., обработку поверхности семян фунгицидами.
В данном случае такая обработка будет эффективной только при использовании системных фунгицидов, проникающих во внутренние ткани семени.
Представители — паразиты культурных злаковых:
I. Род Ustilago, характеризующийся одиночными, сравнительно крупными телио спорами с шиповатой, реже гладкой поверхностью. Около 300 видов этого рода паразитируют на самых разных цветковых растениях. Большинство из них поражает репродуктивные органы.
- 1) U. tritlci — пыльная головня пшеницы;
- 2) U. avenae — пыльная головня овса;
- 3) U. nuda — пыльная головня ячменя;
- 4) U. nigra — черная головня ячменя;
- 5) U. horde! — каменная головня ячменя;
- 6) U. maydis — пузырчатая головня кукурузы.
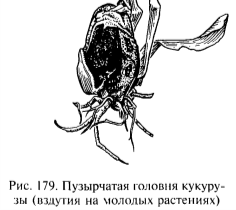
Вызывает местные (локальные) поражения различных органов растения-хозяина: междоузлий, стеблей, листьев, женских и мужских цветков. Заражаться могут и молодые, и взрослые растения, но у взрослых растений восприимчивы только молодые растущие ткани. Здесь дикариотичный мицелий, проникнув в ткани растения-хозяина, разрастается, вызывая местные поражения в виде галлов или вздутий, заполненных телио спорами гриба. Патологический рост этих новообразований обусловлен образуемыми грибом фитогормонами. Мицелий, обильно разрастаясь, разрушает ткань и распадается затем на усто споры. Вначале масса спор прикрыта эпидермисом; после разрыва эпидермиса споры разносятся воздушными течениями. Они способны сразу прорастать, заражая новые растения.
II. Род Sphacelotheca: у этих головневых споровая масса вначале прикрыта оболочкой, состоящей из стерильных гиф и разрушенных тканей хозяина. Позднее оболочка разрывается, а находящиеся под ней телио споры распыляются. Споры одиночные, коричневые или оливковые, чаще с шиповатой поверхностью.
S. panici-miliacei — пыльная головня проса.
III. Род Tilletia. Этот род характеризуется довольно крупными усто спорами, округлыми или овальными, обычно легко распыляющимися. Они образуются чаще всего в завязях пораженных растений. Споровая масса часто имеет селедочный запах (запах триметиламина).
- 1) Твердая (или вонючая) головня пшеницы — средней полосе России вид Т. tritici, в южной — T. foetida;
- 2) Т. controversa — карликовая головня пшеницы.
IV. Род Urocystis. Усто споры соединены в клубочки (спорокучки) по 5- 10 клеток, из которых 1−2 центральные (собственно усто споры), более крупные и более темные, окружены несколькими светлыми стерильными клетками меньшего размера.
Усто споры Urocystis чаще формируются в надземных вегетативных органах растений-хозяев; поражение корней и соцветий встречается гораздо реже. Пораженные части растений искривляются, и на них появляются черные полосы и вздутия. Сначала они покрыты эпидермисом хозяина, позднее разрывающимся.
U. occulta — стеблевая головня ржи.
Заключение
Грибы класса Базидиомицетов человеку необходимо изучать и знать. Они не протяжении всего развития человеческого общества играли важную роль в его хозяйстве, и роль эта двоякая. гриб базидиомицет паразит Подкласс гомобазидиомицеты, к которому относится большинство съедобных грибов, человеку известен лучше, потому что с древних времён люди занимались сбором крупных плодовых тел этих грибов, являющихся относительно легко добываемым питательным блюдом. Немаловажно в данном случае и то, что сельское хозяйство, в частности — посев растительных культур, появилось гораздо позже собирательства, т. е., когда хлеб только-только стали употреблять, грибы (съедобные) уже давно входили в рацион человека. При этом, в связи, грибы собирать не перестали, накопление знаний и дополнение уже имеющихся знаний о них продолжалось, и продолжается сегодня.
Но в связи с активным развитием с/х, растениеводства, из-за огромных площадей посевов и выведения новых сортов появилась необходимость в информации и о вредоносных представителях Базидиомицев. Как видно из приведённых выше данных, для эффективной борьбы с паразитами необходимо знать их жизненные циклы, экологию, физиологию. В виду микроскопичности базидиальных паразитов, а также того факта, что в сознании человека за долгие века понятие «гриб» прочно ассоциировалось со «шляпкой на ножке», углублённое и системное изучение этих паразитических грибов стало возможным лишь с развитием науки, в основном — микробиологии и микологии, а так же биологии в целом.
Поэтому сегодня человек ещё многое не знает о вредных базидиомицетах, но и о полезных, съедобных представителях этого класса знает далеко не всё.