Состав альвеолярного воздуха.
Газовый состав альвеолярного воздуха
Гемоглобин способен захватывать кислород из альвеолярного воздуха (соединение называется оксигемоглобином) и освобождать необходимое количество кислорода в тканях. Особенностью химической реакции кислорода с гемоглобином является то, что количество связанного кислорода ограничено количеством молекул гемоглобина в эритроцитах крови. Молекула гемоглобина имеет 4 места связывания с кислородом… Читать ещё >
Состав альвеолярного воздуха. Газовый состав альвеолярного воздуха (реферат, курсовая, диплом, контрольная)
Газовый состав альвеолярного воздуха обусловлен альвеолярной вентиляцией и скоростью диффузии О2 и СО2 через альвеолярную мембрану. В обычных условиях у человека количество О2, поступающего в единицу времени в альвеолы из атмосферного воздуха, равно количеству О2, диффундирующего из альвеол в кровь легочных капилляров. Равным образом количество СО2, поступающего в альвеолы из венозной крови, равно количеству С О2, которое выводится из альвеол в атмосферу. Поэтому в норме парциальное давление О2 и С О2 в альвеолярном воздухе остается практически постоянным, что поддерживает процесс газообмена между альвеолярным воздухом и кровью капилляров легких.
Таблица 1. Парциальное давление газов в воздушной среде легких.
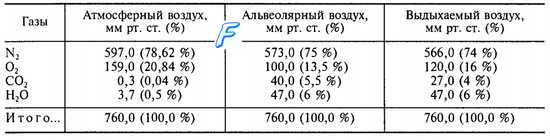
Газовый состав альвеолярного воздуха отличается от атмосферного воздуха тем, что в нем меньше процентное содержание кислорода и выше процент углекислого газа. Состав альвеолярного воздуха отличается от выдыхаемого воздуха большим содержанием углекислого газа и меньшим содержанием кислорода (табл. 1).
Транспорт газов кровью. Транспорт кислорода. Кислородная емкость гемоглобина
Кровообращение выполняет одну из важнейших функций переноса кислорода от легких к тканям, а углекислого газа — от тканей к легким. Потребление кислорода клетками тканей может изменяться в значительных пределах, например при переходе от состояния покоя к физической нагрузке и наоборот. В связи с этим кровь должна обладать большими резервами, необходимыми для увеличения ее способности переносить кислород от легких к тканям, а углекислый газ в обратном направлении.
При 37 С растворимость О2 в жидкости составляет 0,225 мл * л-1 * кПа-1 (0,03 мл/л/мм рт. ст.). В условиях нормального парциального давления кислорода в альвеолярном воздухе, т. е. 13,3 кПа или 100 мм рт.ст., 1 л плазмы крови может переносить только 3 мл О2, что недостаточно для жизнедеятельности организма в целом. В покое в организме человека за минуту потребляется примерно 250 мл кислорода. Чтобы тканям получить такое количество кислорода в физически растворенном состоянии, сердце должно перекачивать за минуту огромное количество крови. В эволюции живых существ проблема транспорта кислорода была более эффективно решена за счет обратимой химической реакции с гемоглобином эритроцитов. Кислород переносится кровью от легких к тканям организма молекулами гемоглобина, которые содержатся в эритроцитах.
Гемоглобин способен захватывать кислород из альвеолярного воздуха (соединение называется оксигемоглобином) и освобождать необходимое количество кислорода в тканях. Особенностью химической реакции кислорода с гемоглобином является то, что количество связанного кислорода ограничено количеством молекул гемоглобина в эритроцитах крови. Молекула гемоглобина имеет 4 места связывания с кислородом, которые взаимодействуют таким образом, что зависимость между парциальным давлением кислорода и количеством переносимого кислорода с кровью имеет S-образную форму, которая носит название кривой насыщения или диссоциации оксигемоглобина. При парциальном давлении кислорода 10 мм рт. ст. насыщение гемоглобина кислородом составляет примерно 10%, а при РО2 30 мм рт. ст. — 50—60%. При дальнейшем увеличении парциального давления кислорода от 40 мм рт. ст. до 60 мм рт. ст. происходит уменьшение крутизны кривой диссоциации оксигемоглобина и процент его насыщения кислородом возрастает в диапазоне от 70—75 до 90% соответственно. Затем кривая диссоциации оксигемоглобина начинает занимать практически горизонтальное положение, поскольку увеличение парциального давления кислорода с 60 до 80 мм. рт. ст. вызывает прирост насыщения гемоглобина кислородом на 6%. В диапазоне от 80 до 100 мм рт. ст. процент образования оксигемоглобина составляет порядка 2. В результате кривая диссоциации оксигемоглобина переходит в горизонтальную линию и процент насыщения гемоглобина кислородом достигает предела, т. е. 100. Насыщение гемоглобина кислородом под влиянием РО2 характеризует своеобразный молекулярный «аппетит» этого соединения к кислороду.
Значительная крутизна кривой насыщения гемоглобина кислородом в диапазоне парциального давления от 20 до 40 мм рт. ст. способствует тому, что в ткани организма значительное количество кислорода может диффундировать из крови в условиях фадиента его парциального давления между кровью и клетками тканей (не менее 20 мм рт. ст.). Незначительный процент насыщения гемоглобина кислородом в диапазоне его парциального давления от 80 до 100 мм рт. ст. способствует тому, что человек без риска снижения насыщения артериальной крови кислородом может перемещаться в диапазоне высот над уровнем моря до 2000 м.
Общие запасы кислорода в организме обусловлены его количеством, находящимся в связанном состоянии с ионами Fe2+ в составе органических молекул гемоглобина эритроцитов и миоглобина мышечных клеток.
Один грамм гемоглобина связывает 1,34 мл О2. Поэтому в норме при концентрации гемоглобина 150 г/л каждые 100 мл крови могут переносить 20,0 мл О2.
Количество 02, которое может связаться с гемоглобином эритроцитов крови при насыщении 100% его количества, называется кислородной емкостью гемоглобина. Другим показателем дыхательной функции крови является содержание О2 в крови (кислородная емкость крови), которое отражает его истинное количество, как связанного с гемоглобином, так и физически растворенного в плазме. Поскольку в норме артериальная кровь насыщена кислородом на 97%, то в 100 мл артериальной крови содержится примерно 19,4 мл О2.