Типы наследования по признаку «высота растений» у гибридов f2 озимого ячменя

Число делянок зависело от количества семян в каждой комбинации. Посев проводился без повторений. Для проведения гибридологического анализа по каждой комбинации высевались родительские формы. Гибриды F2 высевались селекционной сеялкой СКС-6−10. В гибридном питомнике F2 уборка осуществлялась вручную (не менее 300 растений — гибриды; не менее 150 растений — родительские формы), выдергиванием… Читать ещё >
Типы наследования по признаку «высота растений» у гибридов f2 озимого ячменя (реферат, курсовая, диплом, контрольная)
Аннотация
В статье представлены результаты генетического анализа наследования гибридов F2. Рассчитаны степень и частота трансгрессий.
Ключевые слова: озимый ячмень, гибрид, трансгрессия, степень доминирования.
In the article, the results of the genetic analysis of inheritance of hybrids F2 are presented. Degree and frequency of transgression are calculated.
Keywords: winter barley, hybrid, transgression, domination degree.
Селекционная работа по выведению новых сортов зерновых культур является более эффективной, если опирается на информацию о наследовании признаков, получаемую с помощью генетического анализа (Костылев П.И., 2008).
Наличие информации о числе генов, отвечающих за наследование изучаемых признаков, их силы, направления доминирования и взаимодействия между ними позволит ускорить селекционный процесс по созданию новых сортов озимого ячменя, обладающих высоким уровнем продуктивности.
Количественные признаки продуктивности растений определяются полимерными генами и характеризуются широким спектром изменчивости под влиянием окружающей среды. В связи с этим важно знать, как наследуются у гибридов хозяйственно-ценные признаки родительских форм.
Поскольку изменчивость и наследование зависят от генотипа и условий внешней среды, наибольшую ценность представляет информация, полученная в конкретной агроклиматической зоне, для которой создаются новые сорта. Поэтому изучение закономерностей наследования количественных признаков в условиях Ростовской области с целью создания новых сортов является весьма актуальным.
Высота стебля тесно коррелирует с устойчивостью к полеганию. В основном вся западноевропейская селекция основывается на выведении короткостебельных сортов ячменя. Использование генов короткостебельности в селекции ячменя на устойчивость к полеганию является важным мероприятием, хотя требует осторожного подхода. В наибольшей мере этот путь приемлем в селекции сортов в зонах достаточного увлажнения и орошаемого земледелия (Звейнек И.А., 2006).
У ячменя известно несколько типов карликов: брахитичный (brh), «курчавый» (cud), вегетативный (dwf), многоузловый (mnd), «мелкий» (min-en-min), узколистный (nld), uzu, (uz), Bikini, мутанты сложной группы (com), слабые карлики (sld) (Nilan R.A., 1975; Franckowiak J.D., 1999). Все перечисленные мутации рецессивны. Кроме основной характеристики — значительного уменьшения размеров большинства органов, они характеризуются специфическими эффектами, резко снижающими урожайность (Звейнек И.А., 2006).
Описаны новые локусы для 15 генов ячменя, которые влияют на высоту растений (Franckowiak J.D., Lundqvist U., 2002). Они включают 6 мутантов брахитик: brh3 — локализация неизвестна, brh4 — на хромосоме 5HL, brh7 — на хромосоме 7HS и brh8 — на хромосоме 3GS. У трех мутантов из группы слабых карликов были локализованы гены: sld3 на хромосоме 4HS, sld4 на хромосоме 2Н и sld5 на хромосоме 3НS. Выявлено 4 гена (mnd3, mnd4, mnd5, mnd6), контролирующие многоузловость мутантных карликов. Гены mnd3, mnd4 были локализованы ранее на хромосомах 3Н и 5НL соответственно. Полудоминантный ген Ari-s (breviaristatum-s) уменьшает части цветка и укорачивает высоту (Nilan R.A., 1975). Так как существует большая коллекция эректоидных мутантов (ert) и их фенотипы частично похожи на полукарликовые формы, то идентификация новых локусов затруднена. Для уменьшения числа тестов на аллелизм предложены символы генов Zeo, pyr и dsp для большого числа мутантов, уменьшающих расстояние между члениками колосовой оси (Franckowiak J.D., 1999).
Синтетическая селекция основана на использовании для отбора исходного материала, создаваемого путём гибридизации различных сортов и форм с.-х. растений. Трансгрессивная синтетическая селекция — это отбор в расщепляющихся после гибридизации поколениях особей с трансгрессиями и последующая их генетическая стабилизация. На основе синтетической селекции создано подавляющее большинство новых сортов с.-х. растений.
На использовании трансгрессий основан эколого-географический принцип подбора родительских пар — важнейший в современной селекции. Трансгрессии и новообразования у гибридов возникают наиболее часто при использовании этого метода потому, что многие хозяйственно полезные признаки у растений обусловлены несколькими полимерными генами. В результате генетической рекомбинации при скрещивании в отдельных генотипах происходит трансгрессивное сочетание в одном генотипе полимерных генов аддитивного действия, что обусловливает более сильное выражение признака в сравнении с обеими родительскими формами.
Решение проблемы устойчивости к полеганию зерновых культур селекционным путем связывают, прежде всего, с высотой растений. По мнению Н. А. Калашник и др. (2005) даже без учета анатомо-морфологических особенностей соломины вероятность устойчивости низкорослых генотипов выше, чем высокорослых.
Исследования проводились в лаборатории селекции и семеноводства озимого ячменя Всероссийского научно-исследовательского института зерновых культур им. И. Г. Калиненко (ВНИИЗК). В нашем опыте использовались шестирядные сорта озимого ячменя различного эколого-географического происхождения: шестирядные — Мастер, Тигр (ВНИИЗК им. И. Г. Калиненко, г. Зерноград, Россия); Федор (КНИИСХ им. П. П. Лукьяненко, г. Краснодар, Россия); Циндарелла (Saatzucht Josef Breun, Германия) и двурядные — Премьер (КНИИСХ им. П. П. Лукьяненко, г. Краснодар, Россия), Mascara (Secobra Recherches, Франция).
Число делянок зависело от количества семян в каждой комбинации. Посев проводился без повторений. Для проведения гибридологического анализа по каждой комбинации высевались родительские формы. Гибриды F2 высевались селекционной сеялкой СКС-6−10. В гибридном питомнике F2 уборка осуществлялась вручную (не менее 300 растений — гибриды; не менее 150 растений — родительские формы), выдергиванием растений с корнями. Растения с каждой делянки отдельно связывали в снопы и этикировали. На этикетке указывали название питомника, номер делянки и дату уборки.
Опыт закладывался по схеме диаллельных скрещиваний 6×6.
Родительские формы, принимавшие участие в скрещивании, по высоте растений были распределены следующим образом (рис. 1).
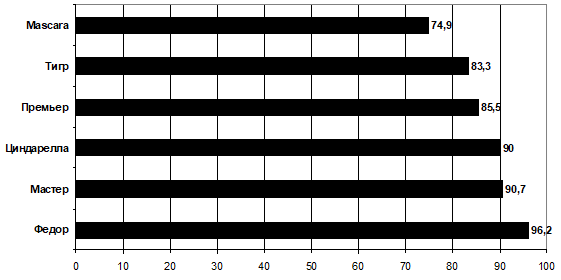
Рис. 1 — Распределение родительских форм по высоте растений
Наиболее приближены друг к другу по изучаемому признаку сорта Мастер и Циндарелла (90,7 и 90,0 см, соответственно). Анализ наследования гибридов, полученных при скрещивании данных родителей, показал, что они, несмотря на близость значений, отличаются по двум парам генов с различным аллельным состоянием (рис. 2).
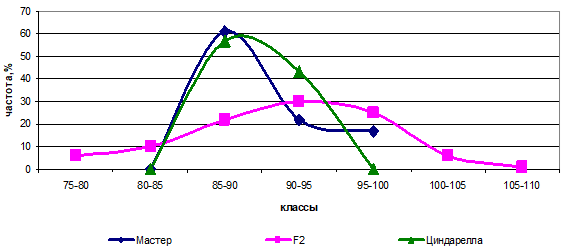
Рис. 2 — Распределение частот родительских форм и гибрида Мастер х Циндарелла по высоте растений
Явным доказательством дигенного расщепления является наличие трансгрессии, что говорит о разнице в аллельном состоянии генов, то есть присутствии доминантных генов, находящихся в различных локусах (ААвв х ааВВ). Расщепление у гибрида Мастер х Циндарелла имеет вид 1:6:9, степень доминирования — 5,86.
Родительские формы Премьер и Циндарелла, имея разницу между собой в 4,5 см, отличались по одной паре генов, что говорит о моногенных различиях (аа х АА). При скрещивании этих сортов расщепление у гибрида составило 1:3 (hр = -0,11; As = -1,11) (рис. 3).
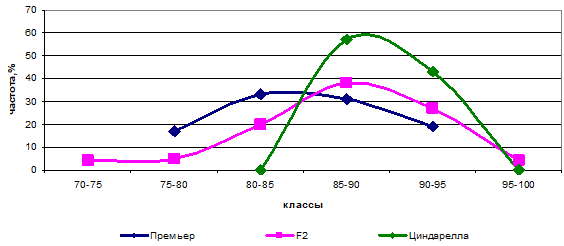
Рис. 3 — Распределение частот родительских форм и гибрида Премьер х Циндарелла по высоте растений
Вершина распределения частот графика располагается в одном классе с вершиной большего родителя (Циндарелла), что говорит о доминировании высокорослости. ячмень гибрид ген короткостебельный Аналогичный тип расщепления был выявлен у комбинации Mascara х Федор, несмотря на то, что родительские формы отличались на 21,3 см.
Мастер и Циндарелла по высоте растений были приблизительно одинаковы (разница — 0,7 см). В комбинации с Федором обе родительские формы различались по двум парам генов с различным аллельным состоянием. В результате скрещивания данных сортов и Федора появились новые генотипы с образованием трансгрессивных форм (рис. 4).
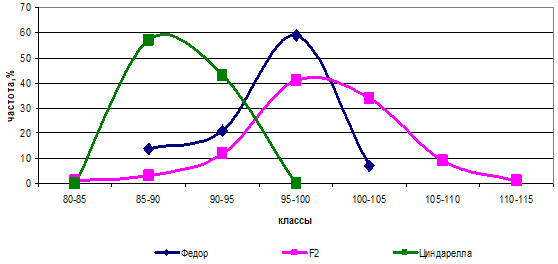
Рис. 4 — Распределение частот родительских форм и гибрида Федор х Циндарелла по высоте растений
Соотношение в комбинации Федор х Циндарелла было 1:15, причем на долю «15» приходились все генотипы кроме гомозигот по рецессивным генам. Гетерозиготные генотипы превысили по высоте генотипы с гомозиготным состоянием доминантных генов и на графике показали положительную трансгрессию. При этом в комбинации Федор х Циндарелла родители различались по генам b и с (ААВВСС х ААввсс), а в комбинации Федор х Мастер — по генам а и с (ААВВСС х ааВВсс) (рис. 5).
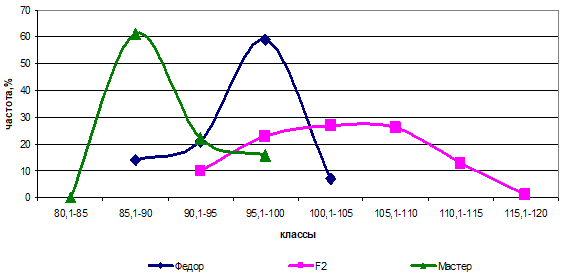
Рис. 5 — Распределение частот родительских форм и гибрида Федор х Мастер по высоте растений
Несмотря на большую разницу по высоте растений между сортами Премьер и Mascara (85,5 и 74,9 см, соответственно) в комбинации с участием данных родителей наблюдались моногенные различия по одной паре генов большой силы, увеличивающей значение гибрида сразу на 11 см. Соотношение классов расщепления — 1:3. Для данного случая может быть приемлема следующая генетическая конфигурация: ааввссDD (Премьер) х ааввссdd (Mascara), где последняя пара генов в доминанте у Премьера влияет на увеличение высоты растений у гибрида.
В комбинации Тигр х Циндарелла наблюдалась дигенная схема расщепления в соотношении 1:6:9, где доле `1' соответствует генотип ааввсс, доле `6' - ааввСС + ААввсс, а доле `9' - А_ввС_. Исходя из этого, можно сделать вывод о том, что родители отличались по локусам А и С (сила гена — 6,7 см) (рис. 6).
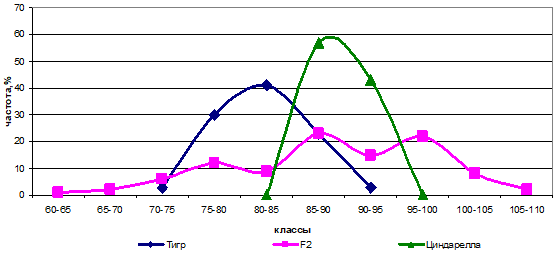
Рис. 6 — Распределение частот родительских форм и гибрида Тигр х Циндарелла по высоте растений
Соотношению 1:15 соответствовала гибридная комбинация Федор х Тигр, что говорит о различиях между родителями по двум парам генов. Отличались данные сорта по локусам А и В (ААВВСС х aabbСС) (рис. 7).
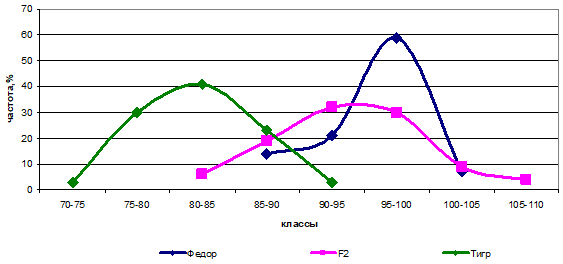
Рис. 7 — Распределение частот родительских форм и гибрида Федор х Тигр по высоте растений
Также по признаку «высота растений» были выявлены такие типы соотношений как 3:1 (Тигр х Федор, Циндарелла х Mascara), 9:7 (Премьер х Мастер, Тигр х Мастер), 7:9 (Циндарелла х Тигр, Мастер х Федор), 1:6:9 (Мастер х Циндарелла).
По нашему мнению, для селекции на устойчивость к полеганию необходимо уделять особое внимание гибридным комбинациям, у которых во втором поколении выщепились отрицательные трансгрессии по высоте растений. Всего было выделено семь подобных гибридов. Степень трансгрессии колебалась от 0,73 (Mascara х Циндарелла) до 0,89 (Циндарелла х Мастер). Наибольшая частота трансгрессии (Тч) была выявлена у комбинаций Циндарелла х Мастер (12%) и Мастер х Премьер (39%). Следует отметить, что чаще всего формы с отрицательной трансгрессией выщеплялись у гибридов, полученных с участием сорта Циндарелла (71,4% комбинаций).
Выводы
- 1. Анализ наследования высоты растений в F2 позволил установить, что данный признак наследуется по-разному в зависимости от конкретной комбинации и величины признака у родителей. Практически у большинства комбинаций (73,3%) доминировала высокорослость.
- 2. Различия по высоте у родительских сортов определялись небольшим количеством генов (1 — 2 парами), что позволяет легко комбинировать их в селекционной работе с генами других количественных признаков.
- 3. Было выделено две комбинации, выщепившие трансгрессивные формы по комплексу хозяйственно-ценных признаков Мастер х Премьер и Мастер х Циндарелла.
- 4. Рекомендуем вовлекать сорт Циндарелла в скрещивания при селекции на низкорослость.
- 1. Звейнек И. А. Некоторые особенности наследования высоты растения у короткостебельного сорта ячменя Golden Promice. / И. А. Звейнек. // Труды по прикладной ботанике, генетике и селекции: «Генетические ресурсы ржи, ячменя и овса». Т. 162. — С.-Петербург, 2006. — С.21−28.
- 2. Калашник Н. А. Генетика продуктивности и показателей качества зерна пивоваренного ячменя в условиях Среднего Прииртышья: Монография. // Н. А. Калашник, Г. Я. Козлова, Н. И. Аниськов. — Новосибирск, 2005. — С. 26, 34, 46, 53.
- 3. Костылев П. И. Генетический анализ количественных признаков риса, сорго и ячменя. // Генетические основы селекции: Материалы Всероссийской школы молодых селекционеров им. С. А. Кунакбаева; 11−15 марта 2008 года. — Уфа: ГНУ БашНИИСХ, 2008. — С. 172−175.
- 4. Franckowiak J.D. Coodinator,s report: Semidwarf genes. / J.D. Franckowiak. — V. 29. — BGN, 1999. — P. 23−31.
- 5. Franckowiak J.D. Descriptions of barley genetic stocks for 2001. // J.D. Franckowiak, U. Lundqvist. — V. 32. — BGN. 2002. — P. 47−137.
- 6. Nilan R.A. Barley, Handbook of genetics. / R.A. Nilan. — V. 2. — 1975. — P. 93−110.