Структурная организация ENaC и топология канала в мембране

ENaC являются важными участниками трансэпителиального транспорта. Каналы данного типа образованы тремя гомологичными субъединицами: б, в и г (Canessa C.M. et al., 1994) (Рис. 3). Позднее были обнаружены еще две субъединицы ENaC: д и е. д-субъединица обладает довольно высокой (37%) идентичностью аминокислотной последовательности по отношению к б-субъединице (Waldmann R. et al., 1995… Читать ещё >
Структурная организация ENaC и топология канала в мембране (реферат, курсовая, диплом, контрольная)
ENaC являются важными участниками трансэпителиального транспорта. Каналы данного типа образованы тремя гомологичными субъединицами: б, в и г (Canessa C.M. et al., 1994) (Рис. 3). Позднее были обнаружены еще две субъединицы ENaC: д и е. д-субъединица обладает довольно высокой (37%) идентичностью аминокислотной последовательности по отношению к б-субъединице (Waldmann R. et al., 1995). Функционально д-субъединица также сходна с б-субъединицей, она может в одиночку образовывать функциональный канал, а также образовывать двг-гетеротример (Waldmann R. et al., 1995). е-субъединица так же как и д-субъединица имеет функциональное сходство с б-субъединицей. Она способна к образованию гетеротримера евг, являющегося менее чувствительным к амилориду, и проявляющим зависимость активности канала от кислотности среды (Wichmann L. et al., 2016). Экспрессией в гетерологичных системах (ооциты Xenopus) было показано, что для возникновения амилорид-зависимых Na+ токов достаточно экспрессии только б-субъединиц ENaC. В свою очередь совместная экспрессия б-субъединиц с в и г субъединицами увеличивает проводимость канала более чем в 100 раз (Canessa C.M. et al., 1994).
Субъединицы ENaC состоят из 632−698 аминокислотных остатков, а их молекулярная масса составляет около 90 кДа (Benos D.J., Stanton B.A., 1999) (Рис. 4).
Каждая субъединица канала состоит из четырех функциональных доменов, различающихся по строению и функциям: цитоплазматический N-конец, два трансмембранных сегмента, экстраклеточная петля и цитоплазматический C-конец (Canessa C.M. et al., 1994). (Рис. 3−5).
В составе субъединиц DEG/ENaC обнаружены участки, консервативные по аминокислотному составу. Одни последовательности характерны для всех представителей данного суперсемейства, другие консервативны только в отдельных ветвях. Данные фрагменты представляют собой важные структурные элементы, необходимые для нормального функционирования канала (Мельницкая А.В. и др., 2006) (Рис. 5).
Цитоплазматический N-конец участвует во многих важных функциях: эндоцитозе, сборке канала, воротных характеристиках, деградации канала (Adams C.M. et al., 1997; Grunder S. et al., 1997; Prince L.S. et al., 1998). При изучении активности ENaC крысы с делециями в N-конце, было обнаружено, что делеция 2−67 аминокислотных остатков в N-конце альфа-субъединицы приводит к снижению эндоцитоза ENaC. Исходя из данных результатов, был сделан вывод о наличии консервативного эндоцитотического мотива в N-конце б-субъединицы ENaC (Chalfant M.L. et al., 1999). Также N-концевой домен играет важную роль в регуляции количества каналов в мембране. Он содержит высоко консервативные остатки лизина, с которыми взаимодействует убиквитинлигаза. Считается, что участок с 47 по 50 аминокислотные остатки (KGDK) являются мотивом, связаным с эндоцитозом, а замены в нем или его делеция приводят к росту амилорид-чувствительных токов за счет увеличения числа каналов в мембране (Staub O. et al., 1997; Chalfant M.L. et al., 1998, 1999). Обнаружено влияние точечных мутаций в N-концевом домене на воротные характеристики ENaC. Показано, что точечные мутации, вызывающие замену высоко консервативных остатков глицина на серин (б-rENaC (G95S), в-rENaC (G37S), г-rENaC (G40S)), резко уменьшают амилорид-чувствительные Na+ токи. Показано, что уменьшение амилорид-чувствительных Na+ токов в данном случае связано с уменьшением вероятности открытого состояния ENaC (Chang S.S. et al., 1996; Grьnder S. et al., 1997; Chalfant M.L. et al., 1999).
Трансмембранные участки M1 и M2 (TM1 и TM2) являются гидрофобными сегментами (Рис. 3−5). Это альфа-спиральные участки богатые остатками положительнозаряженных аминокислот лизина и аргинина. Наибольшая концентрация остатков лизина и аргинина обнаружена на цитоплазматической стороне TM-сегментов (von Heijne, G., 1992). На границе фаз (мембрана/вода) обнаружены остатки ароматических аминокислот триптофана и тирозина, которые, по-видимому, вносят вклад в заякоревание и точное позиционирование TM-сегментов в мембране (Hong H. et al., 2007). Сегмент TM2 содержит в себе множество функциональных участков. Сегмент TM2, по-видимому, является селективным фильтром, воротным механизмом канала и содержит сайт связывания амилорида. На роль воротного механизма претендует участок с 527 по 530 аминокислотные остатки (LLSN). Известно, что замена серина в данном участке на крупные аминокислотные остатки приводит к увеличению вероятности открытого состояния канала (Snyder P.M. et al, 1999; Sheng S. et al., 2001; Kellenberger S., Schild L., 2002).
Экстраклеточная петля — самый крупный функциональный домен белков семейства Deg/EnaC (Рис. 3−5). В нем находится примерно 70% всех аминокислотных остатков белка. Экстраклеточная петля ENaC содержит три крупных консервативных участка CRD I, CRD II, CRD III. Данные участки богаты консервативными остатками цистеина (Рис. 3, 5). Например точечная замена C133 в CRD I участке б-субъединицы ENaC приводит к уменьшению активности канала (фенотип «loss-of-function»), а замены C458S, C472S в участке CRD II б-субъединицы ENaC приводит к уменьшению поверхостной экспрессии канала (Firsov D. et al., 1999). Также в CRD I была обнаружена последовательность WYRFHY, которая считается сайтом связывания амилорида, а мутации в данной последовательности приводили к уменьшению амилорид-чувствительности (Kieber-Emmons T. et al., 1995; Li X.J. et al., 1995). Считается, что данные участки также участвуют в поддержании третичной структуры белка (Firsov D. et al., 1999).
Внутриклеточный C-конец содержит в себе участки связывания с другими белками, сигнальными молекулами и ионами, регулирующими ENaC (Hanukoglu I., Hanukoglu A., 2016). Область после TM2 богата остатками положительнозаряженных аминокислот лизина и аргинина, которые могут взаимодействовать с полярными головками мембранных липидов. Данная область является сайтом связывания фосфотидилинозитолтрифосфата в ви г-субъединицах ENaC (Pochynyuk O. et al., 2005, 2007; Di Paolo G., De Camilli P., 2006). Показано также, что С-конец б-субъединицы ENaC содержит участки фосфорилирования протеинкиназой C, сфингозин-зависимыми протеинкиназами (SDK киназами), казеинкиназой 2 (Volk K.A. et al., 2000; Shi H. et al., 2002; Diakov A., Korbmacher C., 2004; Yang L.-M. et al., 2006). На С-конце также находится участок регуляции ENaC белком — регулятором мембранной проводимости при муковисцидозе (CFTR) (Ji H.L. et al., 2000; Bachhuber T. et al., 2005). Кроме того, данный домен б-субъединицы ENaC содержит сайты связывания с элементами цитоскелета: F-актином и б-спектрином (Rotin D. et al., 1994; Mazzochi C. et al., 2006; Sasaki S. et al., 2014). PY-мотив (PPPXYXXL) (Рис. 5), богатый пролином фрагмент в 65−70 аминокислотных остатков, находящийся в C-конце после сегмента TM2, играет важную роль в убиквитинировании ENaC убиквитинлигазой Nedd4, а нарушения в данном участке приводят к тяжелому наследственному заболеванию — синдрому Лиддла (Schild L. et al., 1996; Staub O. et al., 1996, 1997; Kellenberger S. et al., 1998).
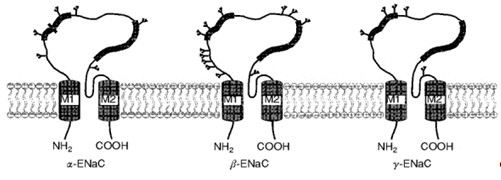
Рис. 3. Топология субъединиц ENaC в мембране. (Модифицировано из Benos, Staton, 1999)

Рис. 4. Аминокислотная последовательность бENaC и топология в мембране. (Модифицировано из Hanukoglu, Hanukoglu, 2016)

Рис. 5. Консервативные домены ENaC. CRD I, II, III — богатые цистеином домены. Deg — экстраклеточный воротный домен. ERD — экстраклеточный регуляторный домен. HG — консервативный His-Gly фрагмент в составе цитоплазматического N-концевого домена. M1, M2 — трансмембранные сегменты. Post-M1 — консервативный участок аминокислотных остатков, следующий за M1. Pre-M2 — участок гидрофобных аминокислотных остатков, находящийся перед M2. PY — высоко консервативный богатый пролином мотив, участок связывания Nedd4. (Модифицировано из Kellenberger, Schild, 2002)