Роль дифференциации в лесных культурах

В лесных культурах по мере полного смыкания крон устанавливается очень тесная связь между размерами стволов и их приростами, благодаря чему ранговое положение (по Е. Л. Маслакову, 1984) стабилизируется и изменяется очень мало. По данным наших исследований, перегруппировка деревьев по степени господства заканчивается в чистых по составу хвойных культурах к 30 годам. Согласно исследованиям Е. Л… Читать ещё >
Роль дифференциации в лесных культурах (реферат, курсовая, диплом, контрольная)
Дифференциация деревьев в культурах начинается на самых ранних этапах становления фитоценоза. Она изначально заложена в самом индивидууме и проявляется еще и в школьных отделениях лесных питомников. Варьирование сеянцев и саженцев по высоте и диаметру стволика в значительной степени определяется различиями генотипа. Попав на лесокультурную площадь и находясь в фазах приживания и индивидуального роста, молодые древесные растения еще больше дифференцируются в результате адаптации к почвенно-грунтовым условиям среды, а затем и в результате социальных отношений.
Биологический прогресс вида по А. Н. Северцову (1939) всегда проявляется в увеличении численности, расселении и распределении на подчиненные группы. Значит, дифференциацию древесных пород в лесных культурах следует считать закономерным биологическим явлением. Этот процесс распределения деревьев по степени господства функционально зависит от густоты посадки и плодородия почвы.
В исследованиях хвойных монокультур зоны смешанных лесов дифференциация деревьев рассматривалась как расчленение их совокупности в зависимости от степени господства индивидуумов. При этом под усилением процесса дифференциации понималось качественное перераспределение деревьев разных классов, выражающееся в уменьшении числа особей высших классов (I—II) и увеличении низших (IV—V). Следовательно, усиление дифференциации сопровождается ухудшением среднего класса Крафта искусственного древостоя в целом. Зависимость процесса дифференциации от первоначальной густоты посадки культур подтверждают данные табл. 8.2, согласно которым отчетливо прослеживается с увеличением густоты уменьшение числа деревьев I— II и увеличение числа деревьев IV—V классов.
Таблица 8.2
Дифференциация в чистых 37-летних культурах сосны в зависимости от густоты посадки.
(свежий бор).
Густота посадки, тыс. шт./га. | Распределение деревьев по классам Крас. | >та, %. | |||
I. | II. | III. | IV. | V. | |
Усиленная дифференциация создает резерв отпада и способствует интенсивному отмиранию деревьев. Наиболее значителен отпад в богатых типах условий произрастания. Так, в культурах сосны с густотой посадки 8 тыс. шт./га к 37-летнему возрасту в свежей сложной субори (С2) он составляет 82%, в свежей простой субори (В2) — 67%, в свежем бору (А2) — 50%. Закономерность этого процесса следует тесно увязывать с первоначальной густотой посадки: она должна быть пониженной в богатых лесорастительных условиях и повышенной в бедных.
Качественная сторона отпада выражается по-разному на различных возрастных этапах. В рядовых культурах ели европейской с густотой посадки 5 тыс. экз./га наблюдается следующая картина: в 17—20 лет (фаза чащи) отмирают экземпляры из III—V классов; в 25—45 лет (фаза жердняка и начало фазы формирования стволов), помимо стволов III—V классов Крафта, в отпад начинают поступать и деревья II класса, а с 55 лет — уже и деревья I класса. В количественном отношении в отпаде культур ели на протяжении 20—55 лет преобладают деревья V класса (до 90%), но по мере увеличения возраста доля представленности деревьев других классов становится все более равномерной. Происходит плавное перераспределение числа стволов отпада в разрезе всех пяти классов, в силу чего в динамике выравнивается средний диаметр элиминирующих стволов со средним диаметром растущих деревьев:
Таблица 8.3
Изменение среднего диаметра древостоев с возрастом.
Показатели. | Возраст, лет. | ||||
Средний диаметр деревьев, см: отпада. | 3,8. | 8,4. | 13,0. | 14,9. | 26,0. |
растущих. | 8,9. | 16,3. | 21,6. | 24,3. | 30,5. |
Разница, %. | |||||
Таким образом, если причиной отпада в древостоях на протяжении двух первых классов возраста можно считать последствия внутривидовой борьбы, то в дальнейшем он все более обуславливается генетической конституцией индивидуумов. В целом дифференциация есть результат совместного проявления наследственных свойств древесных растений и внутривидовой борьбы индивидуумов в древесном сообществе.
Для лесокультурного производства очень важен вопрос о перспективных деревьях, которые должны составлять основу будущего спелого древостоя. В связи с этим интересны данные о стабильности либо, наоборот, изменении во времени особями своего положения в древесном сообществе. Качественный характер перемещений деревьев из класса в класс прослежен нами на примере опытных культур сосны, произрастающих в условиях простой свежей субори (рис. 8.2). Исследования проводили на постоянных пробных площадях с учетом динамики роста каждого живого дерева.
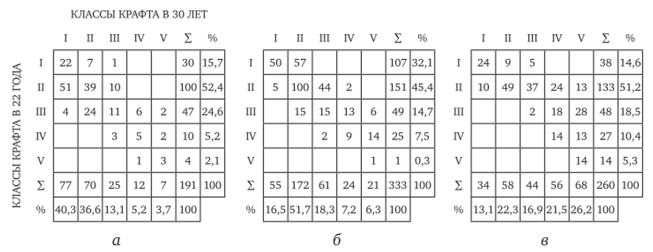
Рис. 8.2. Матрицы дрейфа деревьев сосны по классам Крафта за период 22—30 лет.
Густота посадки: а — 2 тыс.; б — 4 тыс.; в — 8 тыс. экз./га В результате 8-летних наблюдений в фазе жердняка выявлено предопределяющее значение густоты искусственного насаждения в формировании его качественной структуры и перемещениях (дрейфах) деревьев из одного класса в другой. За указанный период лишь 40—52% деревьев сохранили свой класс. Остальные индивидуумы либо улучшили, либо ухудшили свое ранговое положение в древостое. При этом в культурах с редкой посадкой (2 тыс. экз./га) 44% древостоев улучшили класс, а в более изначально загущенных (8 тыс. экз./га) произошел массовый (57%) дрейф деревьев в сторону ухудшения своего ценотического положения. Вне зависимости от густоты первоначальной посадки наиболее стабильными по своему положению в искусственных насаждениях оказались деревья I класса (рис. 8.2). Вероятность того, что особи низших классов с возрастом улучшат ранговый статус, уменьшается на участках с повышенной густотой.
Перегущенность древостоев при одних и тех же почвенно-грунтовых условиях, с одной стороны, катализатор усиления процесса дифференциации, с другой — ингибитор роста и развития популяции в целом. В экстремальных условиях чрезмерная густота может привести даже к распаду одновидового древесного сообщества, что фиксируется обычно в фазе жердняка. Однако в засушливых условиях юго-востока Российской Федерации очень большая густота культур сосны вызывает интенсивную дифференциацию, которая в какой-то мере заменяет разновозрастность, присущую естественным молоднякам, чем обеспечивается высокая засухоустойчивость посадок (Миронов, 1970).
Таким образом, процесс дифференциации является механизмом регуляции численности искусственного дендроценоза посредством качественного изменения его строения. В одновозрастных и однопородных культурах она направлена на имитацию положительных свойств структуры разновозрастного древостоя.
Важное значение имеет вопрос о том, каким образом первоначальное состояние особи, например высота сосны в 5-летнем возрасте, может служить определяющим признаком последующего ее роста? В табл. 8.4 на основании данных Г. Р. Эйтингена (1962) отражено последовательное изменение сосны в зависимости от ее высоты в 5летнем возрасте. Исходным материалом стала группировка 5-летних деревцев по 10-сантиметровым ступеням высоты и дальнейшая фиксация их роста в 6 и 12 лет. Как видно из таблицы, за период с 6 до 12 лет деревья сосны низших ступеней имели почти вдвое лучший рост, чем деревья наивысшей ступени. Сосна низшего ранга увеличила высоту в 6,5 раз, а максимального ранга (степени высоты) — в 3,7 раз. Это свидетельствует, что для низкорослых групп с возрастом характерен ускоренный темп роста, для крупномерных особей — замедляющийся (Г. Р. Эйтинген, 1962).
Таблица 8.4
Зависимость роста сосны от ее высоты в 5-летнем возрасте (выровненные значения).
Ступень высоты в 5 лет, см. | Высота, см, в возрасте, лет. | Разница в высоте за шестилетний период, %. | |
В лесных культурах по мере полного смыкания крон устанавливается очень тесная связь между размерами стволов и их приростами, благодаря чему ранговое положение (по Е. Л. Маслакову, 1984) стабилизируется и изменяется очень мало. По данным наших исследований, перегруппировка деревьев по степени господства заканчивается в чистых по составу хвойных культурах к 30 годам. Согласно исследованиям Е. Л. Маслакова (1984) и С. Н. Сеннова (1984), факт ранжирования и стремления особей к постоянству своего рангового статуса свидетельствует об элементах этологических (поведенческих) аспектов в популяциях древесных растений. Ранжирование в целом способствует сохранению популяцией среднего уровня биологически возможного роста.
Наиболее объективным результатом ростовых процессов является объем ствола. Для лесоводов это весьма информативный показатель. На рис. 8.3 показана динамика редукционных чисел по объему стволов разных классов Крафта. Она подтверждает правильность применения при рубках ухода классического низового метода разреживания. В целях создания быстрорастущих культур необходимо делать ставку на деревья высших рангов развития, ибо, по Н. В. Третьякову (1921), «прирост деревьев есть функция их ранга в древостое». Этот процесс можно начинать с сортировки посадочного материала, а затем переходить к выращиванию особей интенсивного роста посредством равномерного расположения их по лесокультурной площади.
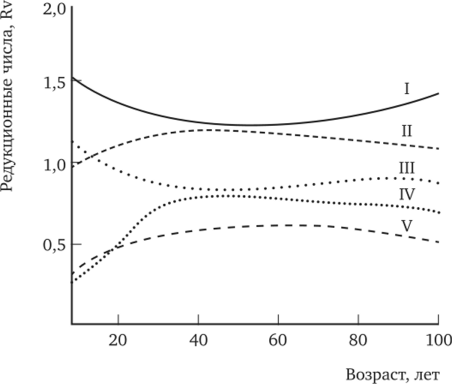
Рис. 8.3. Возрастная динамика редукционных чисел по объему стволов у деревьев разных классов Крафта.
Все сказанное в этой главе относилось к одновозрастным и чистым по составу искусственным насаждениям хвойных пород. Природа их роста во многом адекватна природе естественных одновозрастных хвойных древостоев. Им всем свойственна, как правило, высокая производительность. Возникновение одновозрастных древостоев сосны на гарях — закономерный процесс распространения ее в пределах ареала. В историческом аспекте это явление сопутствовало эволюционному развитию сосны не только как вида, но и как сообщества (одновозрастного древостоя). Чистые по составу одновозрастные искусственные хвойные насаждения надо рассматривать как тип лесных культур, не противоречащий природе естественных древостоев.
Таким образом, дифференциация в лесных культурах — это проявление эволюционно обусловленного процесса, свойственного популяциям древесных растений. Направлена дифференциация на формирование жизнеустойчивого (на протяжении различных фаз роста и развития) древесного сообщества и является средством его численной и качественной саморегуляции. Процесс дифференциации в лесных культурах еще раз подтверждает, что искусственное насаждение следует рассматривать как динамичную (по И. С. Мелехову, 1974) биологическую систему.