Биосинтез ДНК и РНК

К настоящему времени в клетках животных, как и у бактерий, открыто несколько ДНК-полимераз. В репликации ДНК эукариот участвуют два главных типа полимеразпи гВ. Показано, что ДНК-полимераза п состоит из 4 субъединиц и идентична по структуре и свойствам во всех клетках млекопитающих, причем одна из субъединиц оказалась наделенной праймазной активностью. Самая крупная субъединица ДНК-полимеразы… Читать ещё >
Биосинтез ДНК и РНК (реферат, курсовая, диплом, контрольная)
Синтез нуклеиновых кислот ДЫК и РНК из нуклеотидов происходит в ядре клетки и митохондриях.
Синтез ДНК носит название репликация, т. е. создание «дочерних» копийреплик, идентичных «родительской» ДНК.
Синтез РНК носит название транскрипция — «переписывание» информации с матрицы ДНК на матричную РНК (мРНК). Матричная РНК является исходной молекулой для передачи информации о структуре белков, которые синтезируются в плазме. Поэтому ее называют также информационной РНК (иРНК).
В клетке имеется несколько видов РНК. Все они участвуют в синтезе белка. Один из видов РНК носит название транспортной (тРНК). Она осуществляет транспорт аминокислот из цитоплазмы к рибосомальной РНК. Рибосомальная РНК (рРНК) входит в состав рибосом и контролирует биосинтез белка. рРНК имеет более вариабельный состав, чем матричная или транспортная.
Практически все живые организмы, за исключением некоторых видов бактерий, обладают способностью синтезировать нужные им нуклеиновые кислоты.
Матричные, или информационные, РНК синтезируются на одном из участков молекулы ДНК. Такие участки ДНК называют генами. Ген — это участок ДНК, кодирующий информацию о конкретном признаке через последовательность нуклеотидов.
Роль матриц играют разделенные цепи двунитевой материнской ДНК. Репликация представляет собой реакцию полимеризации мононуклеотидов Nub NU2, NU3 и т. д. в дочернюю пол и нуклеотидную цепь (см. разд. 8.5) в последовательности, заданной генами:
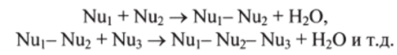
Основную роль в репликации ДНК играет фермент ДНК-полимераза, который соединяет (полимеризует) мононуклеотиды в цепь дочерней ДНК-реплики.
Полимеризационный процесс репликации родительских ДНК включает следующие стадии (рис. 9.27).
- 1. Узнавание места начала процесса в нити ДНК.
- 2. Начало расплетания двойной нити родительских цепей ДНК и образование репликационной вилки.
- 3. Расплетание родительских цепей ДНК в репликационной вилке.
- 4. Начало (инициация) биосинтеза дочерних цепей.
- 5. Рост (элонгация) дочерних цепей.
- 6. Окончание (терминация) процесса репликации ДНК.
Для любого синтеза органических полимеров, осуществляемого in vitro или in vivo, требуется энергия. На реакции полимеризации мононуклеотидов идет энергия, освобождаемая всеми типами дезоксирибонуклсозидтрифосфатов, участвующих в синтезе ДНК. Образующийся пирофосфат РР, под действием пирофосфатазы также расщепляется на две молекулы ортофосфата Р" давая дополнительную энергию для биосинтеза ДНК. После репликации дочерние спирали закручиваются обратно уже без затрат энергии и каких-либо ферментов.
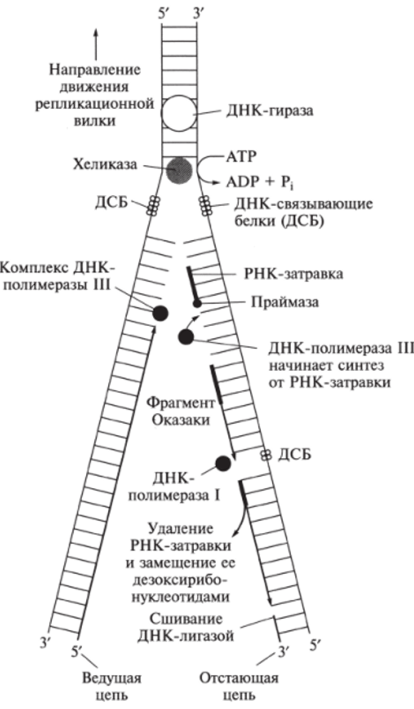
Рис. 9.27. Стадии репликации ДНК (репликативная вилка)
Помимо ДНК-полимеразы в репликации участвует более 40 ферментов и белковых факторов, объединенных в общую ДНК-репликазную систему, называемую реплисомой. Ферменты хеликаза, топоизомераза и ДНК-связывающие белки расплетают ДНК, удерживают матрицу в разведённом состоянии и вращают молекулу ДНК.
Имеется несколько видов ДНК-полимераз, катализирующих репликацию.
Первой была открыта ДНК-полимераза I в кишечной палочке Е. coli. Позже был открыт мутант Е. coli, лишенный ДНК-полимеразы I, но способный синтезировать ДНК с помощью ДНК-полимеразы II. Оказалось, что для репликации ДНК Е. coli необходимо участие нескольких ферментов. ДНК-полимераза I не наделена способностью инициировать синтез цепей ДНК de novo.
Одним из хорошо изученных ферментов, участвующих в стадии инициации репликации ДНК, является специфическая клеточная РНК-полимераза, названная праймазой. Праймаза катализирует синтез короткого олигорибонуклеотида (от 10 до 60 нуклеотидов), т. е. праймера, с которого начинается синтез ДНК. Ираймазы различаются как по структуре, так и по специфичности действия.
Открыт также комплекс белков dna В и dna С, который вблизи репликационной вилки периодически участвует в формировании специфической вторичной структуры ДНК, подходящей для узнавания праймазой.
Основным ферментом, катализирующим биосинтез новообразованной ДНК (точнее, стадию элонгации репликации ДНК), является ДНК-полимераза III, которая представляет собой мультимерный комплекс ДНК-полимеразы (молекулярная масса около 900 000) и ряда других белков. ДНК-полимераза III из Е. coli состоит минимум из 10 субъединиц.
Имеются доказательства, что в димерной форме ДНК-полимераза III катализирует сопряженный синтез ведущей (лидирующей) и отстающей цепей ДНК при репликации. Более точно выяснена также роль ДНК-полимеразы I: она катализирует отщепление затравочного олигорибонуклеотидного праймера и заполнение образующихся после этого пробелов (ниш) дезоксирибонуклеотидами. Известно, что ДНК-полимераза II из Е. coli (молекулярная масса 88 000) выполняет «ремонтные» функции, исправляя повреждения цепей ДНК.
Существенно, что ДНК-полимераза I в качестве матрицы использует одноцепочечные участки, в то время как ДНК-полимераза III — двухцепочечные, в которых имеются короткие одноцепочечные последовательности.
Важную функцию соединения двух цепей ДНК или замыкания двух концов одной цепи ДНК в процессе репликации либо репарации ДНК выполняет особый фермент — ДНК-лигаза, катализирующая за счет энергии АТФ образование фосфодиэфирной связи между ОН-группой дезоксирибозы одной цепи и фосфатной группой другой цепи ДНК.
Функцию раскручивания (расплетения) двойной спирали ДНК в репликационной вилке выполняет специфический белок, названный хеликазой (молекулярная масса 300 000). Одноцепочечные участки ДНК служат в качестве матрицы при репликации. Они стабилизируются при помощи особых белков (ДНК-связывающие белки, молекулярная масса 75 600), препятствующих обратному комплементарному взаимодействию цепей ДНК, в связи с чем эти белки иногда называют дестабилизирующими двойную спираль белками.
Особую роль в с верхе пи рал и заци и играют ферменты топоизомеразы, которые обеспечивают как репликацию, так и транскрипцию ДНК. Эти ферменты наделены способностью не только создавать супервитки, но и уничтожать суперспирализацию путем сшивания образующихся разрывов или разрезания ДНК.
Наконец, открыты специальные ферменты, «редактирующие» ДНК. Они осуществляют вырезание и удаление ошибочно включенных нуклеотидов или устраняют (репарируют) повреждения ДНК, вызванные физическими или химическими факторами (рентгеновское излучение, УФ-лучи, химический мутагенез).
К настоящему времени в клетках животных, как и у бактерий, открыто несколько ДНК-полимераз. В репликации ДНК эукариот участвуют два главных типа полимеразпи гВ. Показано, что ДНК-полимераза п состоит из 4 субъединиц и идентична по структуре и свойствам во всех клетках млекопитающих, причем одна из субъединиц оказалась наделенной праймазной активностью. Самая крупная субъединица ДНК-полимеразы (молекулярная масса 180 000) катализирует реакцию полимеризации, в основном синтез отстающей цепи ДНК, являясь составной частью праймасомы. ДНК-полимераза гВ состоит из 2 субъединиц и преимущественно катализирует синтез ведущей цепи ДНК (см. далее). Открыта также ДНК-полимераза гС, которая в ряде случаев заменяет гВ-фермент, в частности при репарации ДНК (исправление нарушений ДНК, вызванных ошибками репликации или повреждающими агентами).
Следует отметить, что в эукариотических клетках имеется два белковых фактора репликации: RFA и RFC. Фактор репликации RFA связывает одноцепочечную ДНК (наподобие белковых факторов связывания разъединенных цепей ДНК при репликации у Е. coli)y фактор RFC — стабилизатор всего репликационного комплекса.
На первом этапе транскрипции ДНК-полимераза начинает расплетать двойную нить ДНК и образуется репликационная (репликативная) вилка (см. рис. 9.27).
Синтез ДНК в клетках человека и животных происходит в период клеточного цикла, называемого S-фазой.
Репликация ДНК начинается сразу в нескольких тысячах молекул в затравочных участках ДНК — праймерах, синтезируемых специальными ферментами. Затравка синтезируется из рибонуклеозидфосфатов, соответственно ДНК заканчивает синтез новой цепи, дойдя до РНК-затравки. Следствием этого является образование фрагментов Оказаки.
Чтобы обеспечить направленный синтез цепи ДНК, в действие всту пает особая система репараций, удаляющая РНК-затравку путем гидролиза и восстанавливающая правильную струтуру ДНК. Фрагмент завершает ДНК-лигаза, которая соединяет конец 3' нового фрагмента с концом 5' предыдущего.
В ходе транскрипции двойная нить ДНК продолжает расплетаться и вилка перемещается от одного конца молекулы ДНК к другому концу нуклеотид за нуклеотидом: Nu, -> Nu,+j.
Цепи молекулы ДНК расходятся, и каждая из них становится матрицей, на которой синтезируется новая комплементарная цепь. В результате образуются новые двуспиральные молекулы ДНК, идентичные родительской молекуле.
Новые двуспиральные молекулы ДНК состоят из одной цепи исходной родительской молекулы и одной вновь синтезированной цепи.
Состав и последовательность расположения нуклеотидов в родительской ДНК «переписываются» на дочернюю ДНК по тому же принципу комплементарное™, на основе которого образуется двухспиральная молекула ДНК.
Согласно принципу комплементарности свободные нуклеотиды А, Г, Т, Ц (в латинском написании соответственно A, G, Т, С) присоединяются к нити ДНК через водородные связи по схеме А—Т, Г—Ц, Т—А, Ц—Г (рис. 9.28). Напротив каждого нуклеотида одной из родительских цепей ДНК располагается комплементарный нуклеотид синтезируемой ДНК. В результате образующаяся цепочка ДНК представляет собой точную копию исходной цепи.
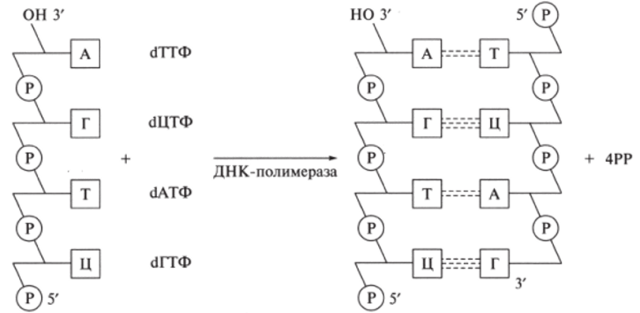
Рис. 9.28. Полимеризация дочерней цепи ДНК по принципу комплементарности
Полимеразы в ходе транскрипции связывают присоединенные нуклеотиды в новую цепь ДНК.
Инициация биосинтеза дочерних цепей ДНК требует предварительного синтеза на матрице ДНК затравочного олигорибонуклеогида — праймера. Праймер имеет свободную гидроксильную группу рибозы. Этот короткоцепочечный олигорибонуклеотид синтезируется комплементарно на матрице ДНК при участии фермента праймазы с полимеразной активностью.
С концевого гидроксила рибозы праймера начинается синтез дочерней цепи ДНК. Синтез осуществляется в результате реакции этерификации между ОНгруппой концевого рибонуклеотида праймера и ОН-группой фосфата первого дезоксирибонуклеотида Nui в строгом соответствии с принципом комилементарности (рис. 9.29). При этом освобождается пирофосфат РР,.
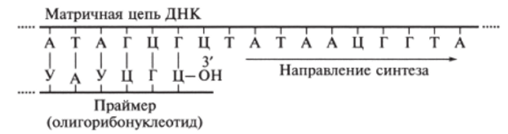
Рис. 9.29. Синтез дочерней цепи ДНК с концевого гидроксила рибозы праймера
В дальнейшем этот фрагмент РНК, комплементарно присоединенный к новообразованной цепи ДНК, разрушается под действием ДНК-полимеразы I и возникшая брешь застраивается олигодезоксирибонуклеотидом при помощи той же ДНК-полимеразы I. Вполне допустимо предположение, что синтез праймера из олигорибонуклеотида имеет глубокий биологический смысл, поскольку в этом случае могут устраняться ошибки, неизбежно возникающие при инициации репликации ДНК.
Предложены различные механизмы биосинтеза ДНК с участием известных ферментов и белковых факторов. Эти механизмы основаны главным образом на данных, полученных в опытах in vitro (в колбе).
На основе изучения кишечной палочки Е. coli в механизме синтеза ДНК выделяют инициацию (начало), элонгацию (продолжение) и терминацию (прекращение) синтеза. Каждая из этих стадий требует участия специфических ферментов и белковых факторов.
Инициация является началом синтеза дочерних нуклеотидных цепей ДНК. В ней участвует как минимум восемь хорошо изученных ферментов и белков. Инициации предшествует синтез праймера на матрице ДНК. В ходе инициации к цепям ДНК последовательно присоединяются ДНК-раскручивающис и ДНКсвязывающие белки, а затем комплексы ДНК-полимераз и праймаз (рис. 9.29).
Инициация представляется единственной стадией репликации ДНК, которая точно регулируется. Однако ее детали не раскрыты и интенсивно исследуются.
Элонгация синтеза ДНК включает два различных по механизму процесса синтеза лидирующей и отстающей цепей на расплетенных родительских цепях ДНК. Лидирующая цепь начинается с синтеза праймера (при участии праймазы) в точке начала репликации. Затем к праймеру присоединяются дезоксирибонуклеотиды под действием ДНК-полимеразы III. Далее синтез протекает непрерывно, следуя за перемещением репликационной вилки.
Синтез отстающей цепи на второй цепи ДЫК протекает в направлении, обратном движению репликационной вилки, и начинается фрагментарно. Фрагменты синтезируются раздельно, начиная с синтеза праймера. Праймер может переноситься с готового фрагмента в точку старта биосинтеза последующего фрагмента при помощи одного из белковых факторов репликации. Элонгация завершается отделением олигорибонуклеотидных праймеров, объединением отдельных фрагментов ДНК при помощи ДНК-лигаз и формированием дочерней цепи ДНК.
Предполагается, что может осуществляться сопряженный и согласованный механизм синтеза лидирующей и отстающей цепей ДНК при участии полимераз и всего комплекса праймаз.
Терминация синтеза ДНК наступает, когда ДНК-матрица пройдена до конца.
Точность репликации ДНК чрезвычайно высока. Возможна одна ошибка на 1000 звеньев цепи, однако и эта ошибка обычно исправляется за счет процессов репарации. Правильность репликации обеспечивается принципом комплементарности и активностью ДНК-полимеразы, способной распознать и исправить ошибку.
Принцип комплементарности, на основе которого построена двухспиральная молекула ДНК, действует и при синтезе иРНК. Гак же как и при синтезе ДНК, с каждым нуклеотидом одной из цепей ДНК связывается водородными евзями комплементарный нуклеотид иРНК. Следует отметить, что в иРНК вместо нуклеотида Т присутствует нуклеотид У. Таким образом, напротив Гднк располагается Црнк" напротив Цднк — Гжк, напротив Аднк — Урнк, напротив ТДНк — Атк.
В результате синтезированная цепь иРНК представляет собой точную копию цепи ДНК. Таким путем информация, содержащаяся в гене, переписывается на иРНК. Поэтому этот процесс и назвали транскрипцией (лат. «transcriptio» — переписывание).
По завершении транскрипции молекулы иРНК перемещаются к месту синтеза белка, т. е. к рибосомам. Туда же из цитоплазмы поступают аминокислоты, из которых строится белок. Каждая аминокислота переносится в рибосому специализированной транспортной РНК (тРНК). Так как в состав природных белков входит 20 разных аминокислот, очевидно, существует не менее 20 разных тРНК.
В цепи тРНК имеются 4−7 последовательных нуклеотидных звеньев, комплементарных друг другу. На рис. 9.30 они обозначены буквами А, Б, В, Г. В этих участках между комплементарными нуклеотидами образуются водородные связи.
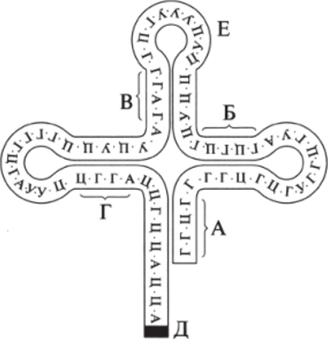
Рис. 9.30. Структура тРНК
Генетический код.
Первое основание. | Второе основание. | Третье основание. | |||
У (А). | Ц (Г). | А (Т). | Г (Ц). | ||
У (А) | Фен | Сер | Тир | Цис | У (А) |
Фен | Сер | Тир | Цис | Ц (Г) | |
Лей | Сер | ; | ; | А (Т) | |
Лей | Сер | ; | Три | Г (Ц) | |
Ц (Г) | Лей | Про | Гис | Apr | У (А) |
Лей | Про | Гис | Apr | Ц (Г) | |
Лей | Про | Глн | Apr | А (Т) | |
Лей | Про | Глн | Apr | Г (Ц) | |
А (Т) | Иле | Тре | Аси | Сер | У (А) |
Иле | Тре | Аси | Сер | Ц (Г) | |
Иле | Тре. | Лиз | Apr | А (Т) | |
Мет | Тре | Лиз | Apr | Г (Ц) | |
Г (Ц) | Вал | Ала | Асп | Гли | У (А) |
Вал | Ала | Асп | Тли | Ц (Г) | |
Вал | Ала | Глу | Гли | А (Т) | |
Вал | Ала | Глу | Г’ли | Г (Ц) | |
Таблица 9.3 в результате возникает сложная петлистая структура, похожая по форме на листок клевера. В верхней части молекулы тРНК (рис. 9.30, Е) расположен триплет нуклеотидов, который по генетическому коду соответствует определенной аминокислоте. Этот триплет называют кодовым триплетом.
У ножки «листка клевера» (рис. 9.30, Д) находится участок, связывающий аминокислоту. Нуклеотидный состав кодовых триплетов тРНК комплементарен нуклеотидному составу триплетов и PH К.
Например, у аланиновой тРНК кодовый триплет ЦГА (в иРНК ему комплементарен триплет ГЦУ). у валиновой тРНК кодовый триплет ЦАА (в иРНК ему комплементарен триплет ГУУ) (табл. 9.3). У тРНК, изображенной на рисунке, кодовый триплет УУУ, соответствующий аминокислоте лизину, — лизиновая тРНК. Она присоединяет и транспортирует в рибосому аминокислоту лизин.
В табл. 9.3 приведен состав триплетов, которыми закодированы все 20 аминокислот (названия аминокислот сокращены).
Пользоваться таблицей просто. Первый нуклеотид в триплете берется из левого вертикального ряда, второй — из верхнего горизонтального и третий — из правого вертикального. Там, где пересекутся линии, идущие от всех трех нуклеотидов, и находится искомая аминокислота. Допустим, нужно узнать, о какой аминокислоте несет информацию триплет УГГ в иРНК. Слева по вертикали берем У, сверху — Г, справа по вертикали — Г. Линии пересекаются на «Три», т. е. триптофан. В ДНК эта аминокислота закодирована триплетом АЦЦ.
Синтез дочерней молекулы дезоксирибонуклеиновой кислоты происходит в процессе деления клетки на матрице родительской молекулы ДНК. При этом генетический материал, зашифрованный в ДНК, удваивается и делится между дочерними клетками. В клеточном цикле время синтеза и удвоения ДНК называется S-стадией. За ней следуют стадии G2 и М — митоз (деление клетки).