Щитовидная железа.
Эндокринная и центральная нервная системы, высшая нервная деятельность, анализаторы, этология

Парафолликулярные клетки (К-клетки) располагаются в интерфолликулярных островках и стенке фолликулов щитовидной железы. Они существенно отличаются по своей структуре от тиреоцитов: имеют низкую электронную плотность (светлые клетки) и содержат в цитоплазме большое количество секреторных гранул и везикул. К-клетки не контактируют с коллоидом и максимально приближены к капиллярам… Читать ещё >
Щитовидная железа. Эндокринная и центральная нервная системы, высшая нервная деятельность, анализаторы, этология (реферат, курсовая, диплом, контрольная)
Щитовидная железа — одна из самых крупных эндокринных желез, располагается в нижней части гортани и верхней части трахеи в виде двух лопастей, соединенных между собой перешейком, лежащим на вентральной поверхности трахеи. Снаружи покрыта соединительнотканной капсулой, в толще которой располагаются мелкие сосуды, капилляры, а также миелинизированные и немиелинизированные нервные волокна. От капсулы внутрь железы отходят соединительнотканные тяжи (септы), по которым проходят кровеносные и лимфатические сосуды, нервные волокна. Паренхима органа включает в себя несколько типов клеток: тиреоидные.
(формирующие стенку фолликула), парафолликулярные и интерфолликулярные.
Фолликулярный эпителий состоит в основном из паренхимных клеток и образует основную структурно-функциональную единицу щитовидной железы — фолликул (округлой или овальной формы пузырек). Между фолликулами находятся прослойки соединительной ткани, пронизанные кровеносными капиллярами. Просвет фолликула заполнен коллоидом, который непосредственно прилежит к апикальной поверхности тиреоцита с многочисленными микроворсинками, образованными выступами плазматической мембраны. Противоположная сторона тиреоцита тесно контактирует с перикапиллярным пространством, причем в базальном отделе клетки плазматическая мембрана образует глубокие складки, что значительно увеличивает ее поверхность, контактирующую с кровеносной системой. Боковые поверхности тиреоцитов соединяются за счет замыкательных комплексов, что надежно изолирует внутреннее пространство фолликула от окружающей соединительной ткани. В цитоплазме тиреоцитов особенно сильно развит шероховатый эндоплазматический ретикулум из многочисленных канальцев и более крупных полостей, заполненных гомогенным мелкозернистым материалом. Комплекс Гольджи очень хорошо выражен и представлен тремя основными компонентами: крупными вакуолями, уплощенными цистернами (ламеллами) и микропузырьками. В зоне комплекса Гольджи обнаруживаются секреторные гранулы различной величины, электронной плотности и формы, окруженные мембраной. Кроме характерных гранул в апикальной части цитоплазмы выявляются коллоидные капли, окруженные мембраной.
Исходными продуктами для биосинтеза йодированных гормонов служат аминокислота тиронин и йодид. Поставляемый с кровью йодид в тиреоцитах, подвергаясь окислению, превращается в молекулярный йод, который вступает в соединение с тиронином, и через ряд промежуточных соединений (монойодтиронин, дийодтиронин) образуются тироксин (тетрайодтиронин) и трийодтиронин. Считается, что эти гормоны синтезируются в свободном состоянии в цитоплазме фолликулярных клеток и необходимое для организма количество гормона сразу после образования поступает в кровь, а его избыток включается в молекулы тиреоглобулина и накапливается в коллоиде. Тиреоглобулин — крупная гликопротеиновая молекула, синтезированная по обшей схеме синтеза белка. Освобождение тиреоглобулина из секреторных гранул в полость фолликула происходит либо путем диффузии через ограничивающую мембрану, либо путем экзоцитоза. Иодирование тиреоглобулина происходит в основном в просвете фолликула и на границе между коллоидом и апикальной плазматической мембраной. При снижении потребности организма в тиреоидных гормонах внутриклеточный гидролиз тиреоглобулина прекращается и белок выводится в полость фолликула, где накапливается в виде гормонального резерва. Накопленный в полости фолликула тиреоглобулин поступает в цитоплазму тиреоцита за счет микропиноцитоза коллоида: при повышенной потребности возможно захватывание крупных порций с помощью псевдоподий. Образовавшиеся в результате эндоцитоза коллоидные капли постепенно перемещаются к базальной части клетки; при встрече с лизосомами происходят их тесный контакт и слияние, причем гидролитические ферменты лизосом поступают внутрь коллоидных капель. Коллоидные капли уменьшаются, а поступившие в цитоплазму тиреоидные гормоны диффундируют в перикапиллярное пространство и затем проникают в кровеносное русло. При истощении запасов коллоида в полости фолликула синтез, йодирование и протеолитическое расщепление тиреоглобулина полностью осуществляются в цитоплазме тиреоцитов (рис. 12.9).
Трийодтиронин непрочно связан с белками крови и быстро проникает в клетки, а у тироксина эта связь прочнее, поэтому он способен длительно циркулировать в кровеносном русле. Физиологическим действием в основном обладает трийодтиронин, так как он в 5…6 раз активнее тироксина и, проникая в клетки тканеймишеней, воздействует на ферментные системы, локализованные в митохондриях (ферменты трикарбонового цикла, фосфорилирования, железосодержащие дыхательные и др.). Гормоны шитовидной железы регулируют процессы роста и развития, величину энергетического обмена, белковый, углеводный, жировой, водный и минеральный обмен, тем самым влияя на деятельность сердца, нервную систему и половые железы.
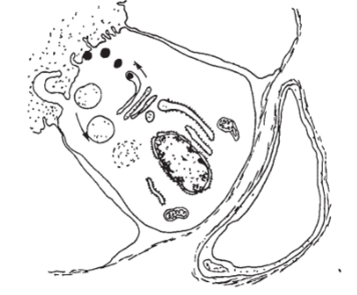
Рис. 12.9. Схема секреторного процесса в тиреоците.
Демонстративным примером участия тиреоидных гормонов в регуляции роста и развития служит влияние щитовидной железы на метаморфоз амфибий. При удалении щитовидной железы головастики никогда не превратятся во взрослую форму, вырастая, остаются на стадии личинки, а скармливание щитовидной железы или введение на ее основе препаратов ускоряют метаморфоз амблистомы (личиночной формы аксолотлей) и других земноводных. У птиц и млекопитающих гормоны щитовидной железы играют важную роль во время эмбрионального развития. При выключении функции железы у птиц тормозится рост тела, задерживаются процессы окостенения в конечностях, нарушается процесс эмбрионального оперения и т. д. С активностью щитовидной железы связаны и видовые особенности развития птиц. Так, у выводковых видов птенцы сразу после вылупления из яйца способны бегать и самостоятельно питаться, так как щитовидная железа активно функционирует уже на ранних стадиях зародышевого развития. У птенцовых видов, птенцы которых вылупляются слепыми и беспомощными, щитовидная железа в эмбриональный период развита слабо и ее активность возрастает только после вылупления из яйца.
Аналогичная закономерность обнаруживается и у млекопитающих — зрелорожденные детеныши (способные самостоятельно передвигаться) имеют щитовидную железу, активно функционирующую на ранних стадиях эмбрионального развития. У млекопитающих, детеныши которых появляются на свет слепыми и голыми, деятельность щитовидной железы проявляется незадолго до рождения. Следует, однако, учитывать, что некоторое количество тиреоидных гормонов потомство получает из организма матери через плаценту и с молоком. Гормоны щитовидной железы существенно влияют на развитие костной ткани. При удалении щитовидной железы за счет действия соматотропного гормона усиливается рост трубчатых костей, стимулируется пролиферация эпифизарных хрящей, но задерживается дифференциация костной ткани.
Введение
тироксина гипофизэктомированным животным вызывает окостенение эпифизов при незначительном росте костей. Рост и прорезывание зубов у млекопитающих зависят от содержания в крови тиреоидных гормонов. Тиреоидные гормоны стимулируют регенерацию тканей и способствуют заживлению ран: грануляционная ткань быстрее заполняет дно раны, и эпителизация идет более интенсивно.
Влияние щитовидной железы ярко проявляется при оценке основного обмена: тиреоидэктомия снижает его на 45…59%. Повышение содержания гормонов за счет введения тироксина увеличивает уровень основного обмена до 50% по сравнению с нормой и выход энергии при выполнении механической работы. Гормоны щитовидной железы резко усиливают окислительные процессы и теплопродукцию, причем этот эффект обнаруживается и на изолированных органах. Инкубируемые фрагменты мышц, печени и почек от животных, которым предварительно вводили тироксин, потребляют значительно больше кислорода, чем аналогичные ткани, взятые у тиреоидэктомированных животных. In vivo при гипофункции щитовидной железы у животных нарушается терморегуляция, понижается температура тела и они плохо переносят охлаждение.
Гормоны шитовидной железы способны значительно повысить потребность организма в метаболитах: усиливается расходование белка и жира, азотистый баланс становится резко отрицательным, азот в составе мочевины выводится с мочой. При гиперфункции щитовидной железы возрастает интенсивность основного обмена: обнаруживается резкое похудание, расходуется до 70% запасов жировых депо, содержание холестерина в крови снижается. Углеводный обмен также интенсифицируется: уровень глюкозы в крови несколько повышается, а печень быстро теряет запасы гликогена. В основе физиологического действия тиреоидных гормонов лежит регуляция интенсивности дыхания клеток, их непосредственное влияние на поглощение кислорода митохондриями и другими структурными компонентами клеток, усиление окислительных реакций и последующее изменение активности ряда ферментов и влияние на ядерный аппарат клеток. Тироксин способствует переносу аминокислот к рибосомам, причем максимальное использование резорбированных аминокислот происходит в том случае, когда все они присутствуют в клетке в соответствии с требованиями определенного количественного соотношения и последовательности информационной РНК.
Развитие и функциональное состояние нервной системы находятся под постоянным влиянием щитовидной железы. Ее удаление приводит к нарушению развития мозга: нейроны имеют меньшие размеры, задерживается миелинизация нервных волокон. Полноценная дифференцировка развивающегося мозга происходит при достаточном уровне тиреоидных гормонов в организме. В ходе дифференцировки и созревания нервной клетки существует критический период, во время которого гормоны оказывают свое влияние на морфогенез, поэтому нарушение функций щитовидной железы в раннем возрасте приводит к кретинизму. В дальнейшем они необходимы для поддержания метаболизма и функциональной активности нейрона. Тиреоидэктомия резко снижает возбудимость животных и ослабляет у них активные и пассивные оборонительные реакции, подавляет деятельность высших нервных центров. При гипотериозе условные рефлексы вырабатываются с большим трудом и требуют постоянного подкрепления: введение тиреоидина восстанавливает рефлекторную деятельность мозга.
Гормоны щитовидной железы существенно влияют на деятельность сердца, изменяя ритм его сокращений. При удалении щитовидной железы сокращения сердца урежаются, а при введении тироксина интактным животным частота сердечных сокращений повышается в 1,5 раза. Передозировка гормона может привести к трепетанию предсердий. При функциональных гипотиреоидных состояниях, характерных для зимнеспящих животных (медведей, ежей, сурков, летучих мышей), наблюдается снижение температуры тела, уменьшение уровня основного обмена и существенное замедление ритма сердечных сокращений. Периоду пробуждения у этих животных предшествует активизация деятельности щитовидной железы; введением тироксина можно прекратить зимнюю спячку и обеспечить пробуждение у ежей. Физиологический гипертиреоз отмечается при функциональных состояниях животных, которые требуют интенсификации обмена веществ: беременность, лактация (особенно у высокопродуктивных животных).
Тиреоидные гормоны ускоряют всасывание глюкозы в желудочно-кишечном тракте, участвуют в регуляции уровня гликогена в печени, усиливают секрецию молока и увеличивают содержание в нем жира. Под действием тиреоидных гормонов резко ускоряются процессы окисления жирных кислот в митохондриях.
Регуляция секреции тиреоидных гормонов осуществляется прежде всего за счет выделения специфического стимулирующего вещества — тиреотропного гормона передней доли гипофиза. У гипофизэктомированных животных уменьшаются размер и масса щитовидной железы, развивается атрофия ткани и в первую очередь редуцируются новообразованные фолликулы на фоне симптомов тиреоидной недостаточности. Повышение секреции тиреотропного гормона увеличивает продукцию и поступление в кровь гормонов щитовидной железы. Но увеличение концентрации тиреоидных гормонов сказывается на продукции тиролиберина в гипоталамусе: избыток гормона тормозит тиреотропную функцию гипофиза, что ведет к снижению секреции тироксина в щитовидной железе и реализации обратной отрицательной связи, обеспечивающей в норме постоянство гормонального уровня.
Наряду с системой гуморальной регуляции, в которой тиреотропная функция гипофиза коррелируется с выделением АКТГ и соматотропного гормона, активность щитовидной железы существенно зависит от функционального состояния и энергетических потребностей организма. При охлаждении, вызывающем раздражение холодовых рецепторов, рефлекторно усиливается секреция тиреотропных гормонов, обусловленная воздействием гипоталамического тиролиберина. Вместе с влиянием гипоталамуса регулирующее воздействие на состояние щитовидной железы оказывает вегетативная нервная система: симпатический отдел усиливает активность органа, а парасимпатический угнетает. Влиянием симпатического отдела вегетативной нервной системы объясняется ускоренное выделение тиреоидных гормонов в условиях стрессовых ситуаций, что обеспечивает приспособление организма к неблагоприятным факторам среды: охлаждению, отрицательным эмоциям и т. д.
Наиболее часто нарушение деятельности щитовидной железы вызвано недостаточным поступлением йода в организм. Снижение уровня тиреоидных гормонов сказывается на деятельности гипоталамуса и гипофиза, который отвечает усиленной секрецией тиреотропина, вызывающего разрастание ткани щитовидной железы (зоб). Увеличение железы может быть за счет разрастания соединительной ткани (простой зоб) или увеличения объема фолликулярной ткани (коллоидный зоб). В местностях, где почва и вода бедны йодом (горные районы, зоны подзолистых, серых лесных и солонцовых почв), возможно развитие зобных эндемий.
Недостаточное поступление йода в организм приводит к снижению образования тиреоглобулина, обладающего свойствами каротиназы (фермента, ответственного за превращение каротина в витамин А). Таким образом, избыток тиреоидных гормонов в тканях организма нейтрализуется витамином, А и в случаях гипертиреоза возможен А-авитаминоз. Поступление витамина, А или каротина с кормом нормализует основной обмен и поднимает уровень гликогена в печени. На активность щитовидной железы оказывают влияние полноценность белков, жиров, углеводов и наличие антитиреоидных веществ гойтрогенов (содержатся в соевых бобах, горохе, белом клевере, капусте и турнепсе), которые избирательно нарушают процессы связывания йода в щитовидной железе. Недостаток тирозина и фенилаланина, избыток растительных жиров и длительное, однообразное углеводное питание приводят к развитию дистрофических изменений в железе и нарушению продукции тиреоидных гормонов.
Парафолликулярные клетки (К-клетки) располагаются в интерфолликулярных островках и стенке фолликулов щитовидной железы. Они существенно отличаются по своей структуре от тиреоцитов: имеют низкую электронную плотность (светлые клетки) и содержат в цитоплазме большое количество секреторных гранул и везикул. К-клетки не контактируют с коллоидом и максимально приближены к капиллярам в интерфолликулярных островках; располагаются на периферии и на наружной поверхности фолликулов. Гранулы, локализованные в цитоплазме К-клеток, имеют диаметр 200…280 нм, содержат тонкогранулированный материал и окружены одиночной мембраной. Вблизи плазматической мембраны обнаруживаются признаки экзоцитоза — слияния мембраны секреторной гранулы с плазмалеммой и освобождения ее содержимого в пери капиллярное пространство. Установлено, что К-клетки продуцируют тиреокальцитонин — полипептид, состоящий из 32 аминокислот, вызывающий падение уровня кальция и фосфора в крови. Это снижение концентрации кальция в циркулирующей крови обусловлено торможением его мобилизации из костей, причем существенно меняется деятельность клеток костной ткани: угнетаются остеокласты и активируются остеобласты, что приводит к фиксации кальция и фосфора. Наряду с этим существенно увеличивается удаление фосфора с мочой. Тиреокальцитонин усиленно секретируется и поступает в кровь при повышении концентрации кальция, что является важным звеном в поддержании постоянства его уровня во внутренней среде организма.
Околощитовидные (паращитовидные) железы. У млекопитающих представляют собой парные образования и расположены около щитовидной или вилочковой железы либо погружены в их ткань. Эти небольшие округлые железы окружены собственной соединительнотканной капсулой. От капсулы в толщу органа отходят тонкие септы, между которыми находятся тяжи и группы железистых клеток — паратиреоцитов. Паратиреоциты располагаются радиально вокруг многочисленных капилляров, пронизывающих тело органа. Структурная организация клеток паренхимы околощитовидной железы носит характерные признаки интенсивного секреторного процесса: развитой эндоплазматический ретикулум представлен многочисленными уплощенными канальцами и свободнолежащими рибосомами, обусловливающими мелкозернистую структуру цитоплазмы; комплекс Гольджи хорошо развит и представлен большим количеством микропузырьков, имеющих мелкозернистое содержимое.
Кроме микропузырьков в цитоплазме паратиреоцитов обнаруживаются более крупные (диаметр 150…200 нм) гранулы, формирующиеся в комплексе Гольджи и выделяющиеся из клеток в области латеральных клеточных контактов. Наряду с секреторными структурами паратиреоциты могут содержать липидные капли, липопигментные тельца и лизосомальные образования. Кроме этих клеток, описываемых при световой микроскопии как «главные», паренхима околошитовидных желез содержит электроноплотные, или «оксифильные», клетки, функциональное значение которых еще не установлено. Не исключена возможность того, что околощитовидные железы образованы клетками одного типа (главными клетками), цитоплазма которых может меняться в широких пределах в зависимости от функционального состояния. Паратиреоциты контактируют с одним или несколькими капиллярами, причем периэндотелиальное пространство расширено и содержит кроме двух базальных мембран и специальные клетки сосудистого окружения — перициты. Эндотелий капилляров уплощен и имеет многочисленные фенестры.
Если у животного удалить околощитовидные железы (паратиреоидэктомия), то на 2…3-е сутки нарушается возбудимость нервной системы: походка становится более напряженной, появляется фибриллярное подергивание отдельных мышц головы и туловища. Единичные сокращения усиливаются и переходят в бурные приступы общих судорог (титания), которые повторяются все чаще. Животное вскоре погибает за счет понижения содержания кальция в крови, поскольку определенный уровень кальция необходим для течения важнейших жизненных процессов. Внутривенное введение хлористого кальция оперированным животным способно временно прекратить судороги и снизить возбудимость ЦНС.
Околошитовидные железы секретируют гормон — паратгормон, полипептид с молекулярной массой около 8500, состоящий из 83 аминокислотных остатков. Паратгормон регулирует концентрацию кальция и фосфора. В результате его действия повышается уровень кальция в крови и тканях организма, а содержание фосфатов снижается. Исходя из того, что кальций попадает в организм с пищей и поступает в кровь в тонком кишечнике, действие гормона проявляется уже на уровне кальцийтранспортирующих систем — энтероцитов. Основным депо кальция в организме служит костная ткань, содержащая до 99 % всего кальция в составе гидрооксиапатита.
Действие паратгормона проявляется в мобилизации запасенного в кости кальция. Усиливается деятельность разрушающих кость остеокластов и подавляется функция фиксирующих кальций остеобластов, происходит деполимеризация мукополисахаридов основного вещества кости, ее декальцинация и поступление ионов кальция и фосфорной кислоты в кровь. Одновременно с этим уменьшается реабсорбция фосфатов из мочи: они быстро выводятся, и их уровень в крови быстро снижается. Продукция паратгормона регулируется в основном уровнем кальция в крови по принципу обратной отрицательной связи. В случаях высокой потребности организма в кальции, наступающей при беременности и лактации, когда для развивающегося плода или образования казеина необходимо поступление дополнительных значительных количеств кальция, обнаруживается физиологическая гиперфункция околощитовидной железы. Если не увеличить содержание кальция в корме, то возможна декальцинация (остеомаляция и остеопороз) костной ткани материнского организма.