Особенности пищеварения в кишечнике жвачных

Это предположение подтверждается экспериментами по выяснению влияния частичного удаления поджелудочного сока и желчи на процессы пищеварения и молокообразования. Вне опыта поджелудочный сок и желчь через фистульную трубку поступали в двенадцатиперстную кишку. При относительно длительном времени лишения животных определенного количества пищеварительных соков удалось проследить влияние дефицита… Читать ещё >
Особенности пищеварения в кишечнике жвачных (реферат, курсовая, диплом, контрольная)
ПИЩЕВАРЕНИЕ В ТОНКОМ КИШЕЧНИКЕ
В тонком кишечнике совершаются основные биохимические процессы переваривания и всасывания. Пищеварение в тонком кишечнике обеспечивает расщепление сложных питательных веществ до тех форм (в основном мономеров), в которых они могут всосаться из кишечника в кровь и лимфу. В кишечнике происходит окончательный гидролиз пищевых веществ под действием секретов поджелудочной железы, кишечного сока и желчи и всасывание продуктов гидролиза. К тонкому отделу кишечника относят двенадцатиперстную, тонкую и подвздошную кишки. В полость двенадцатиперстной кишки открываются протоки поджелудочной железы и желчный проток. Натощак содержимое двенадцатиперстной кишки имеет слабощелочную реакцию (pH 7,2−8,0). При переходе в нее порций желудочного содержимого реакция среды становится кислой, однако быстро происходит ее нейтрализация за счет желчи, поджелудочного сока и кишечного сока. Эти секреты прекращают действие желудочного пепсина. Чем выше кислотность дуоденального содержимого, тем больше выделяется поджелудочного сока и желчи.
Ритм моторики кишечника не зависит от моторики сычуга. В связи с тем, что панкреатический и желчный протоки впадают в кишечник на значительном удалении от пилоруса, в начальной части двенадцатиперстной кишки сохраняется кислая реакция и продолжаются характерные для сычуга процессы (рис. 8).
Рис. 8.
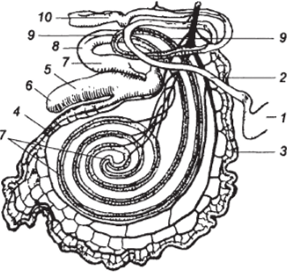
Кишечник крупного рогатого скота.
1 — пилорус желудка; 2 — двенадцатиперстная кишка; 3 — тощая кишка; 4 — подвздошная кишка; 5 — слепая кишка и ее б — верхушка; 7 — ободочная кишка и ее б — начальная; 9 — конечная петля; 10 — прямая кишка.
Поджелудочный сок и его значение. Роль поджелудочной железы в пищеварительном процессе определяется ее внешнесекреторной деятельностью, то есть выделением поджелудочного сока в просвет двенадцатиперстной кишки. Количество выделяемого в сутки поджелудочного сока в среднем составляет у жвачных 6−7 л.
Поджелудочный сок представляет собой бесцветную жидкость щелочной реакции, которая обусловливается высокой концентрацией анионов НСО3. Кроме того, в поджелудочном соке присутствуют хлористый натрий, хлористый кальций, фосфаты и другие неорганические ионы.
В поджелудочном соке содержатся ферменты, действующие на все группы питательных веществ: протеолитические, гликолитические (амилолитические) и липолитические.
К протеолитическим ферментам относятся:
11 трипсин — основной протеолитический фермент поджелудочного сока. Трипсин первоначально выделяется в своей неактивной форме — в форме трипсиногена. Последний активируется под действием фермента, вырабатываемого слизистой оболочкой двенадцатиперстной кишки — энтеропептидазы (энтерокиназы). Трипсин активен в щелочной среде. Этот фермент гидролизует белки и их промежуточные соединения — альбумозы и пептоны — до полипептидов, дипептидов и даже аминокислот;
- 11 химотрипсин — также выделяется в неактивном состоянии в виде химотрипсиногена и активируется трипсином. Химотрипсин расщепляет белки преимущественно после их обработки пепсином и трипсином;
- 1! панкреатопептидаза (эластаза) — осуществляет гидролиз специфических белков соединительной ткани и мукополисахаридов, расщепляя их на пептиды и аминокислоты;
- Ш карбоксипептидаза — отщепляет от пептидов свободные аминокислоты со стороны карбоксильной группы;
- 11 дезоксирибонуклеаза и рибонуклеаза — осуществляют гидролиз нуклеиновых кислот;
- Ш липаза поджелудочного сока обладает широким диапазоном действия на жиры, расщепляя их на глицерин и жирные кислоты. Этот фермент растворяется в воде и действует на жиры только на границе раздела вода-жир. Липаза активируется ионами кальция и желчными кислотами.
К гликолитическим (или амилолитическим) ферментам поджелудочного сока относятся:
- Ш амилаза — расщепляет крахмал, гликоген и амилопектин на декстрины и мальтозу;
- Ш глюкозидаза (мальтаза) — расщепляет мальтозу на две молекулы глюкозы;
® фруктофуронидаза — расщепляет сахарозу на глюкозу и фруктозу.
У молодняка в молочный период в поджелудочном соке отмечается высокое содержание галактозидазы, расщепляющей молочный сахар — лактозу на глюкозу и галактозу.
Изучением секреции поджелудочного сока и отделения желчи занимались в прошлом многие исследователи (Бернар, 1856, 1859; Бернштейн, 1869; Ландау, 1873; Гейденгайн, 1875; Лебедев, 1876 и др.). В 1888 г. Павлов установил роль блуждающего нерва в секреции поджелудочной железы. Несколько позднее Метт (1889) путем раздражения блуждающего нерва электрическим током подтвердил данные Павлова. Он показал, что раздражение блуждающего нерва увеличивает количество белкового фермента в поджелудочном соке. При раздражении блуждающего нерва электрическим током наряду с усилением секреции отмечалось и ее снижение.
Считается, что секреция поджелудочного сока при действии соляной кислоты происходит рефлекторно и «нервный центр» заложен интрамурально в области привратника желудка. В регуляции секреторной деятельности поджелудочной железы принимает участие симпатическая нервная система. При раздражении симпатического нерва панкреатического сока отделяется больше, чем при раздражении блуждающего. Однако в последнем случае увеличивалось количество ферментов и плотного остатка. Установлено, что в период расщепления жира в двенадцатиперстной кишке количество сока увеличивается.
Введение
атропина в период расщепления жира в кишечнике не снижает секрецию поджелудочной железы, но уменьшает количество сухого остатка и азота. В составе блуждающего нерва показано наличие как секреторных, так и тормозящих волокон. При этом малые дозы атропина почти не влияли на тормозящие волокна блуждающего нерва, в то время как большие дозы снижали секрецию сока.
Существование секретина в дальнейшем было подтверждено многими исследователями. Участие симпатической нервной системы в гуморальной фазе секреции поджелудочной железы подтверждают данные о том, что новокаиновая блокада чревных нервов и симпатических стволов снижает секрецию поджелудочного сока, отделяемого в ответ на введение секретина и соляной кислоты, в 2−3 раза. При этом изменяется и химический состав сока, в частности, понижается щелочность и амилолитическая активность и, наоборот, повышается протеолитическая сила сока. При введении соляной кислоты в двенадцатиперстную кишку после двусторонней перерезки симпатических веток секреция поджелудочного сока снижается. Значительное уменьшение объема сока сопровождается снижением количества плотного остатка и концентрации ферментов. Таким образом, к деятельности поджелудочной железы имеют отношение как парасимпатическая, так и симпатическая системы. Павлов указывал, что поджелудочная железа благодаря нервной регуляции на каждый вид пищи выделяет сок соответствующего количества и качества. Полагают, что поджелудочная железа к роду пищевых режимов не приспосабливается и в отличие от слюнных и желудочных желез регулируется преимущественно гуморальным механизмом при слабом участии нервного фактора. И это является, видимо, основным моментом, объясняющим отсутствие изменений в деятельности поджелудочной железы при качественно различном питании. Возбудителем поджелудочной железы является вода и овощные соки, причем влияние последних более сильное. Сокогонное действие на поджелудочную железу оказывают также жир и продукты его расщепления.
Минеральная вода, введенная через фистулу двенадцатиперстной кишки собак натощак, понижает секрецию поджелудочного сока, в то время как та же вода, но введенная за 5−10 мин до еды, повышает сокоотделение на пищевой раздражитель. При изучении влияния высокой температуры и инсоляции на ферментовыделительную функцию поджелудочной железы показано, что при перегревании организма концентрация ферментов в соке возрастает.
При содержании животных на солнце количество поджелудочного сока и протеолитического фермента в нем становится значительно меньшим, чем в условиях оптимальной температуры.
Опыты на телятах с хронической фистулой панкреатической железы показали наличие непрерывной секреции поджелудочного сока и постоянное содержание в нем белкового, углеводного и жирового ферментов. Они же впервые доказали влияние кормового рациона на непрерывную секрецию поджелудочной железы у сельскохозяйственных животных. Замена в рационе отрубей овсянкой или сена силосом увеличивает переваривающую силу сока на белок. И, наоборот, замена отрубей соломой понижала протеолитическую активность сока. Скармливание животным меляссы повышало концентрацию белкового фермента. В опытах на быке с фистулами сычуга, двенадцатиперстной кишки и поджелудочной железы показано, что соляная кислота и олеиново-кислый натрий при введении их в двенадцатиперстную кишку усиливают секрецию панкреатического сока.
В дальнейшем на свиньях была разработана методика наложения хронической фистулы на проток поджелудочной железы. Преимущество этой методики состоит в том, что вне опыта поджелудочный сок через систему трубок поступает в полость двенадцатиперстной кишки и участвует в пищеварении.
Разработана и методика наложения хронической фистулы на проток поджелудочной железы овец. Поджелудочный сок у них выделяется непрерывно и представляет слегка мутноватую жидкость со слабокислой реакцией (pH 6,5) и содержанием значительного количества сухого вещества. Суточное количество поджелудочного сока у овец колеблется в пределах 240−290 мл и зависит от состава рациона. Наибольшее количество сока отделяется при содержании овец на рационе, состоящем из сена и жмыхов. Наименьшая величина выделяемого сока отмечается при скармливании животным травы. У овец наиболее высокий уровень секреции поджелудочной железы наблюдается в дневные часы и наименьший — в ночное время. Жвачка усиливает секрецию поджелудочного сока. Секреция натощак выше, чем после кормления. Зимой поджелудочного сока отделяется меньше, чем летом.
В первые месяцы жизни ягнят, когда основная часть рациона состоит из молока, протеолитическая активность химуса двенадцатиперстной кишки очень высока. На этом уровне она находится до 3−4-месячного возраста ягнят, затем идет на снижение и к 7−8 месяцам достигает величины, характерной для взрослых животных. Исследования, проведенные на ягнятах с хронической фистулой общего протока поджелудочной железы, показали, что количество и качество поджелудочного сока и желчи с возрастом животного заметно изменяются и изменения имеют определенную закономерность.
Изучение изменений в поджелудочном сокоотделении у телят показало, что количество поджелудочного сока у телят с возрастом увеличивается и достигает к 6 месяцам 1400 мл в сутки, параллельно с этим отмечено снижение переваривающей силы сока. Кроме того, выявлено, что секреция поджелудочного сока у телят в течение суток также неравномерна и зависит от возраста животных и характера кормления. Молоко, подогретое до 45−50°С, увеличивало амилолитическую активность сока и снижало его протеолитическую способность. Концентрированные корма для телят являются сильным раздражителем деятельности поджелудочной железы.
У сельскохозяйственных животных поджелудочная железа отделяет сок непрерывно, даже в период длительного голодания.
Желчеобразование и желчевыделение. Секреция желчи осуществляется клетками печеночной паренхимы, собранными в печеночные дольки. От печеночных клеток начинаются желчные капилляры, которые собираются в желчные протоки. Последние, сливаясь между собой, образуют печеночный проток, открывающийся в желчный пузырь. У лошади, верблюда, оленя желчный пузырь отсутствует, его функцию выполняют хорошо развитые желчные ходы. Общее количество выделяющейся желчи в сутки составляет у лошади и жвачных до б л, у овец — 300−400 мл.
В образовании желчи основная роль принадлежит трем процессам:
Ш активной секреции, например, билирубина, желчных кислот;
Ш фильтрации из плазмы крови воды и растворенных в ней веществ — глюкозы, креатинина, ионов калия, натрия, хлора;
11 реабсорбции — всасывания обратно в кровь некоторых элементов первичной или печеночной желчи. Таким образом, часть веществ, входящих в состав желчи, синтезируются клетками печени, другие поступают в нее из плазмы крови.
В желчном пузыре происходит реабсорбция из желчи некоторых солей и воды, в результате чего из печеночной желчи образуется более густая, концентрированная, так называемая пузырная желчь. Плотность печеночной желчи 1,009−1,013, она содержит 1−5% плотных веществ, pH 7,5. Пузырная желчь содержит 9−20% плотных веществ, плотность ее 1,026−1,048, pH 6,8. Кроме того, в ее состав входит слизь, выделяющаяся бокаловидными клетками слизистой оболочки желчного пузыря. Образование желчи в печени идет непрерывно, однако сфинктер желчного протока находится в сокращенном состоянии, пока в полость кишечника не поступит пищевая кашица.
В состав желчи входят вода, слизь и пигменты (билирубин и биливердин), желчные кислоты (холевая, дезоксихолевая, литохолевая), желчнокислые соли, холестерин и лецитин, соли неорганических кислот. Ферментов в желчи не содержится.
Поступление желчи в кишечник облегчает пищеварение за счет щелочной реакции, нейтрализующей кислый химус, поступающий из желудка. Натриевые соли желчных кислот уменьшают поверхностное натяжение воды и жира, способствуя эмульгированию жиров пищевой кашицы, при этом их поверхность увеличивается и облегчается процесс взаимодействия с липолитическими ферментами. Кроме того, соли желчных кислот активируют липазу поджелудочного сока и ускоряют всасывание конечных продуктов расщепления жира. Желчные кислоты усиливают моторику кишечника. Они соединяются с нерастворимыми в воде жирными кислотами, в результате чего образуются водорастворимые комплексы, всасывающиеся в кишечнике.
В желчи овец содержится до 35 мг% сахара. Внутривенное введение глюкозы увеличивает количество сахара в желчи. Установлено, что с ростом и развитием ягнят и телят, с переходом от молочного питания на растительные корма и повышением потребления кормов количество желчи увеличивается, возрастает щелочность ее и изменяются другие показатели химического состава. Летом секреция желчи выше, чем зимой, но высокая температура и инсоляция, наоборот, угнетают желчеобразовательную функцию печени и снижают концентрацию желчных кислот. На высоте 1850 м над ур. м. при температуре воздуха от +10 до +24°С секреция желчи на пищевые раздражители снижается. Желчеобразование в клетках печени является непрерывным процессом, однако оно может быть стимулировано актом еды, раздражением блуждающего нерва или гуморальным путем, например действием секретина.
Первые порции желчи поступают в кишечник через 5−10 мин после приема пищи. Для накопления и выделения желчи важна синхронная работа мышц сфинктера желчного протока и мускулатуры желчного пузыря. При эвакуации желчи сокращение стенок пузыря сопровождается расслаблением сфинктера. При накоплении желчи наблюдается обратная картина — расслабление мускулатуры стенок пузыря и сокращение сфинктера. Эта согласованная работа мышц регулируется нервнорефлекторным и гуморальным путем. Раздражение блуждающего нерва приводит к сокращению мышц пузыря и расслаблению круговой мышцы сфинктера, т. е. эвакуации желчи.
Стимуляция симпатического нерва прекращает поступление желчи в кишечник. Гуморальная регуляция выделения желчи связана с выработкой в слизистой оболочке двенадцатиперстной кишки гормона холецистокинина. Его образование стимулируют промежуточные продукты гидролиза белков и жиров. Данный гормон вызывает сокращение мышц пузыря и расслабление мышцы сфинктера желчного протока. Важным гуморальным возбудителем желчевыделения является сама желчь. Поступление желчи в кишечник способствует желчеобразованию.
Таким образом, желчеотделительная функция печени, как и секреторная деятельность поджелудочной железы, изменяется в зависимости от характера кормления, возраста животного и температуры окружающей среды.
Эксперименты показали, что секреторная активность поджелудочной железы и желчеотделение меняются в зависимости от физиологического состояния животного. Полученные данные свидетельствуют, что по ходу суягности изменяется как количество желчно-панкреатического сока, так и его качество. Причем эти изменения становятся более заметными начиная со второй половины беременности животных.
После установления определенного фона в деятельности поджелудочной железы и желчеотделения у беременных животных приступают к основному опыту — выяснению особенности секреции желчно-панкреатического сока в период лактации. С наступлением лактации среднесуточное количество желчно-панкреатического сока составило 2525 мл. Сок имел pH 6,8 ± 0,12.
Следовательно, секреторная деятельность поджелудочной железы и печени во втором месяце лактации находится на уровне предыдущего месяца. Заметное уменьшение секреции желчно-панкреатического сока наблюдается в третьем месяце лактации — 2103 ± 103,83 мл в сутки — с содержанием плотного вещества 4,65 ± 0,19%. Переваривающая сила на белок (3,4 ± 0,062 ед.), на углеводы (266 ± 0,054 см3) и на жиры (5,4 ± 0,763 см3) остается на исходном уровне; pH сока смещается в щелочную сторону и равен 7,05 ± 0,065. Значительное изменение в секреторной деятельности поджелудочной железы и желчеотделительной функции печени происходит в последнем месяце лактации. Так, количество желчно-панкреатического сока снижается до 1955 ± 91,165 мл с содержанием плотного вещества 4,39 ± 124%. В связи с уменьшением количества желчно-панкреатического сока повышается переваривающая сила его на белок (3,8 ± 0,206 ед.), увеличивается количество углеводов (80 ± 35,751 см3) и жиров (5,0 ± 0,671 см3), pH сока равен 7,04 ± 0,07.
Если сравнить количество желчно-панкреатического сока, отделившегося в период лактации, с характером секреции во время суягности и после отъема ягнят, то видно, что максимальная секреция сока падает на период лактации. При этом наибольшее количество сока выделяется в первый период. Со второй половины лактации интенсивность отделения сока постепенно снижается. Если нарастание количества его по ходу лактации выразить в процентном отношении, то в первом месяце лактации объем сока увеличивается на 54% по сравнению с исходным уровнем. Во втором месяце лактации это повышение составляет 40, в третьем — 20%. В последнем месяце лактации секреторная активность поджелудочной железы и печени превышает контрольную величину на 11%. Из этих данных видно, что к концу лактационного периода секреция сока резко снижается, в то время как концентрация плотного вещества и ферментов колеблется в пределах исходных величин, а в большинстве случаев имеет тенденцию к увеличению. Однако, принимая во внимание обильную секрецию поджелудочного сока и желчи в начальный период лактации, можно заметить, что наибольшее количество ферментов выделяется в первой половине лактационного периода.
Так, например, самое высокое число ферментных единиц наблюдается в первом месяце лактации и составляет: на белок — 8702 ± 648,6, углеводы — 616 118± 89 215и жиры — 13 705 ± 2293,52. Количество плотного вещества в суточном объеме сока равно 118,22 ± 8,15 г. Во втором месяце лактации отмеченные показатели желчно-панкреатического сока незначительно снижаются и составляют: белкового фермента — 8690 ± 470 ед., углеводного — 572 779 ± 70 746 и жирового — 11 357 ± 2198 см3. В суточном количестве сока содержится 112,55 ± 5,15 г плотного вещества. В третьем месяце лактации количество ферментов в желчно-панкреатическом соке меньше, чем в предыдущем месяце, и составило соответственно 7437 ± 422,0, 550,242 ± 52 660 и 10 741 ± 2364 см3. Содержание плотного вещества в данном случае также ниже — 99,65 ± 7,62 г. В последнем месяце лактации с желчно-панкреатическим соком ферментов выделяется столько же, сколько выделилось их в третьем месяце лактационного периода, и составляет: для белка — 7429 ± 412,0 ед., углеводов — 547 400 ± 50 210 и жиров — 9775 ± 1311 см3.
Увеличение количества желчно-панкреатического сока и повышение его ферментативной активности в период лактации связано, по-видимому, с физиологической перестройкой обмена веществ в организме, вызванной процессом молокообразования. Об этом свидетельствуют показатели изменения суточных удоев и химического состава молока у подопытных животных по ходу лактации.
В первом месяце лактации среднесуточный удой у овцематок составил 1136 ± 71,50 мл, с содержанием жира 5,53 ± 0,383% и белка — 4,15 ± 0,112%; в переводе на суточную продукцию: жира — 62,821 и белка —47,144 г. В третьем месяце лактации суточный удой уменьшается до 758 мл. Значительное снижение количества молока наблюдается в последнем, четвертом, месяце лактации и равняется 508 ± 71,26 мл в сутки. При этом в молоке содержится: жира — 7,72 ± 0,516%, белка — 5,70 ± 54%, что в переводе на суточную продукцию составляет ответственно 39,217 и 28,956 г.
При сравнении секреции желчно-панкреатического сока с суточным удоем подопытных животных на протяжении всей лактации между ними обнаруживается определенная динамическая связь. Так, интенсивная секреция поджелудочного сока и желчи наблюдается в первом месяце лактации и совпадает с наибольшим суточным удоем и высокими показателями химического состава молока. Затем, по ходу лактации, по мере снижения молочности, отмечается уменьшение количества желчно-панкреатического сока. Наименьший объем сока и молока устанавливается в последнем месяце лактации.
Отмеченная связь между секреторной функцией поджелудочной железы и печени, с одной стороны, и деятельностью молочной железы, — с другой, у лактирующего животного возникла не случайно, а развивалась постепенно в ходе эволюции и в своей основе имеет биологический смысл. Последний состоит, по-видимому, в том, что обилие желчно-панкреатического сока с высокой переваривающей силой ускоряет расщепление большего количества корма и улучшает усвоение его организмом.
Это предположение подтверждается экспериментами по выяснению влияния частичного удаления поджелудочного сока и желчи на процессы пищеварения и молокообразования. Вне опыта поджелудочный сок и желчь через фистульную трубку поступали в двенадцатиперстную кишку. При относительно длительном времени лишения животных определенного количества пищеварительных соков удалось проследить влияние дефицита их на организм. В контрольном опыте было установлено, что у лактирующих коров в сутки выделяется 4−4,3 л поджелудочного сока и 16 л желчи. Наибольшее количество поджелудочного сока и желчи выделяется в дневные часы, во время и после кормления животных. Наименьшая секреция пищеварительных соков наблюдается ночью, когда животные отдыхают. После установления фона сокоотделения у подопытных животных удаляли некоторое количество поджелудочного сока. Наблюдения показали, что исключение из процессов пищеварения 50−70% от общего количества поджелудочного сока, почти не отражаясь на состоянии животных, снижало жирность молока с 4,5 до 3,7%. При этом отмечалось незначительное уменьшение суточного удоя.
Более глубокое изменение в организме подопытных животных происходило при исключении из процессов пищеварения желчи. Следует отметить, что при удалении 50% желчи в рубцовом пищеварении и общем состоянии животных резких изменений не отмечалось: незначительно повысилась общая кислотность среды рубцового содержимого, соотношение кислот осталось неизменным (уксусной — 60−70, пропионовой — 16−19 и масляной — 12−14%).
Надой молока снизился на 30−33% и находился на таком уровне 3 дня после восстановления поступления желчи. Значительно снизилась жирность молока — в среднем на 1,5%. Плотность и кислотность молочного жира колебались в пределах нормы. Заметно отклонился показатель Рейхерта — Мейселя: он снизился с 26 до 22,80. При исключении в течение суток 70% выделившейся желчи общее состояние животных было несколько угнетено. Нарушилось пищеварение в желудочно-кишечном канале, удой молока снижался на 40−50% и восстанавливался только на 4−5-й день. Наиболее низкий удой приходился на 2−3-й день после начала опыта. Жирность молока снижалась с 4,5−5 до 3,3% и восстанавливалась в течение 4 дней после окончания опыта. Пищеварение в рубце существенно не изменилось, претерпевало изменение только соотношение кислот. Так, концентрация уксусной кислоты на следующий день после опыта повысилась с 60 до 70 м экв/л, а пропионовой — снизилась с 14 до 10 м • экв/л.
Как видно из приведенных данных, при исключении части указанных пищеварительных соков наступало довольно стойкое и глубокое нарушение процессов молокои жирообразования. Кроме того, было парализовано нормальное пищеварение в кишечнике, в результате большое количество питательных веществ корма, по-видимому, не использовалось организмом. При этом надо учесть, что при исключении желчно-панкреатического сока пищеварение в преджелудках не подвергалось заметному изменению. Микробиологические и биохимические процессы в рубце во время опытных дней протекали на уровне исходных величин, характерных для нормы. Эти данные еще раз свидетельствуют о том, что поджелудочный сок и желчь имеют важное значение для процесса пищеварения и обмена веществ и в конечном итоге оказывают большое влияние на продуктивность животного.
Наибольшему уровню отделения желчно-панкреатического сока в период лактации соответствует наивысший суточный удой и максимальное содержание жира и белка в молоке. Таким образом, выявляется связь между секрецией желчно-панкреатического сока с секрецией молока. Чем выше секреторная деятельность поджелудочной железы и печени, тем больше молочная продуктивность животного.
Глубокое и длительное снижение молочной продуктивности наблюдалось при исключении из процессов пищеварения смешанного сока (поджелудочный сок 4- желчь). Наряду с указанными изменениями у животных отмечено полное расстройство деятельности желудочно-кишечного канала и некоторое нарушение в обменных процессах. Уменьшился щелочной резерв по Неведову и снизилась концентрация сахара в крови.
Данные, полученные в экспериментах на лактирующих животных, показывают, что с наступлением лактации также увеличиваются секреторная деятельность околоушных слюнных и желудочных желез и желчеотделение. Причем наибольшее количество пищеварительных соков выделяется в первой половине лактации и зависит от уровня молочной продуктивности. Чем выше молочная продуктивность животного, тем больше выделяется пищеварительных соков с высокой переваривающей силой. Со второй половины лактационного периода интенсивность отделения соков снижается до минимума к моменту запуска.
Усиление деятельности пищеварительных желез у лактирующего животного, по всей вероятности, связано с повышением обмена веществ. У лактирующих животных потребность в кормах увеличивается, в связи с чем повышается и деятельность органов пищеварения. Усиление функции пищеварительной системы у лактирующих животных связано не с обильным кормлением, а с повышением деятельности всего организма. Наибольший уровень кровообращения, газообмена и дыхания у коров наблюдается после отела и совпадает с вершиной лактационной кривой. Значительное повышение деятельности дыхательной, сердечно-сосудистой систем отмечено у коров-рекордисток.
Определенные взаимоотношения отмечались между химическими компонентами пищеварительных соков и составной частью молока. Наибольшее количество пищеварительных ферментов и наивысшая суточная продукция молочного жира и белка отмечались в первой половине лактации. Начиная со второй ее половины вышеуказанные показатели постепенно снижались и достигали наименьшей величины к моменту запуска.
Следовательно, путем повышения секреторной функций пищеварительных желез можно увеличить скорость переваривания корма и количество используемых организмом питательных веществ. Согласно учению Павлова (1897), обильное отделение сока с самого начала еды есть наилучшее условие для появления аппетита, от которого в значительной степени зависит перевариваемость и усвояемость пищи. Наибольшее отделение пищеварительных соков совпадает с периодом наивысшей продуктивности — с наступлением лактации усиливается деятельность пищеварительной системы, повышается степень использования кормов, а значит, увеличивается и продуктивность животных. Снижение секреции соков наблюдалось при охлаждении поверхности вымени и воздействии высокой температуры. Следовательно, воздействие неадекватных раздражителей, сопровождающееся болевой реакцией, по-видимому, приводит к торможению секреторной деятельности пищеварительных желез.
Таким образом, в период лактации у животных с рецепторов молочной железы постоянно поступают импульсы, которые усиливают или ослабляют деятельность пищеварительной системы. Специальной серией опытов установлено, что взаимосвязь между пищеварительными и молочными железами осуществляется с помощью рефлекторного механизма. Причем степень проявления рефлекса зависит от вида и индивидуальных особенностей животного. У коз рефлекторное отделение пищеварительных соков в ответ на раздражение молочной железы происходит более активно и с меньшим латентным периодом, чем у овцематок. Следовательно, раздражение рецепторов молочной железы, связанное с процессом доения, является не только стимулом для усиления функции молочной железы, но и оказывает определенное влияние на деятельность пищеварительных систем. Это имеет особо важное значение для лактирующего организма, особенно в период раздоя животного.
В период раздоя животного все процессы, связанные с подготовкой молочной железы к дойке, и акт доения вызывают усиление секреции пищеварительных соков. Последние способствуют появлению аппетита, в результате повышается поедаемость корма, а в итоге увеличивается продуктивность животного. Из практики раздоя коров можно привести множество примеров, когда при сочетании обильного кормления с подготовкой молочной железы к доению добивались высоких удоев.
Дальнейшие исследования показали, что импульсы, возникающие при раздражении рецепторов молочной железы во время дойки, массажа вымени и т. д., передаются по афферентным нервам молочной железы в центральную нервную систему, а затем по эфферентным нервам достигают органов пищеварения. В данном случае афферентные волокна проходят в составе наружного семенного нерва, в дорзальных и боковых канатиках спинного мозга. Эфферентными нервами пищеварительной системы являются блуждающий нерв и секреторные волокна чревных нервов. Переключение афферентного пути на эфферентный происходит на разных уровнях ЦНС. По предположению Грачева (1964), это может происходить на уровне продолговатого, среднего и промежуточного мозга, подкорковых образований и в коре больших полушарий. По нашим данным, место замыкания рефлекторных дуг пищевых реакций, возникающих при раздражении рецепторов молочной железы, находится в передних и средних ядрах гипоталамуса. Не исключена возможность участия в указанной рефлекторной дуге других отделов ЦНС.
Таким образом, установлено, что у животных в период лактации с молочной железы постоянно поступают импульсы, которые усиливают или ослабляют деятельность пищеварительной системы. Адекватные раздражители, которые обычно применяются в практике (дойка, массаж, обмывание и обтирание вымени), способствуют повышению деятельности пищеварительных желез.
Неблагоприятные воздействия на организм животных, такие как чрезмерное нагревание поверхности вымени или высокое внутривыменное давление, тормозят секрецию пищеварительных соков. В том и другом случае влияние осуществляется с помощью нервного механизма.
В настоящее время показано, что в регуляции деятельности пищеварительных и молочных желез наряду с нервным фактором участвует эндокринная система, в частности щитовидная железа, надпочечники и половые железы. Гипофункция щитовидной железы вызывает снижение секреции пищеварительных соков, удоя и особенно содержания жира в молоке.
Кроме щитовидной железы, на деятельность пищеварительной системы и молочной железы оказывают влияние надпочечники. Выключение хромаффинной ткани надпочечников снижает секрецию пищеварительных желез и содержание ферментов в соках. При введении адреналина адреналэктомированным животным сокоотделение восстанавливается до исходного уровня. У этих животных снижались удой и содержание жира в молоке, а введение адреналина восстанавливало их до нормы. В регуляции деятельности пищеварительных желез у животных определенную роль играют и половые железы. Хорошо известна тормозящая роль половых гормонов, обеспечивающих подавление пищеварительной функции в период течки и охоты.
После удаления яичников секреция пищеварительных соков у овец и коз временно снижалась и восстанавливалась через 60 дней после кастрации. У этих животных не отмечалось посткастрационного увеличения сокоотделения, характерного для кастрированных плотоядных животных.