Тотипотентность ядра соматической клетки

В качестве генетического маркера, гарантировавшего чистоту эксперимента, было использовано число ядрышек. Лягушки, от которых брали ядра, образовывали два ядрышка на ядро, т. е. каждый ядрышковый организатор гомологичных хромосом функционировал нормально. В качестве донора соматических ядер использовали X. 1аеу1 В, гетерозиготных по делении ядрышкового организатора, поэтому имевших только одно… Читать ещё >
Тотипотентность ядра соматической клетки (реферат, курсовая, диплом, контрольная)
Дж. Гердон продемонстрировал возможность полного развития Хепорш !aevis на основе генетической информации ядра соматической клетки. Неоплодотворенные яйца X. 1аеу1л облучали большими дозами ультрафиолетового света и таким образом убивали их ядра. Затем в энуклеированное яйцо инъецировали ядро из эпителия кишечника головастика. В ряде случаев из таких яиц развились головастики, а затем — взрослые лягушки (рис. 10.1).
В качестве генетического маркера, гарантировавшего чистоту эксперимента, было использовано число ядрышек. Лягушки, от которых брали ядра, образовывали два ядрышка на ядро, т. е. каждый ядрышковый организатор гомологичных хромосом функционировал нормально. В качестве донора соматических ядер использовали X. 1аеу1 В, гетерозиготных по делении ядрышкового организатора, поэтому имевших только одно ядрышко на ядро. Все лягушки, развившиеся в результате пересадки ядер, имели по одному ядрышку.
Таким образом, эксперименты показали, что дифференцировка клеток в онтогенезе не обязательно сопровождается необратимой инактивацией генетического материала ядра, а проблема генетического контроля индивидуального развития тесно связана с проблемой дифференциальной экспрессии генов.
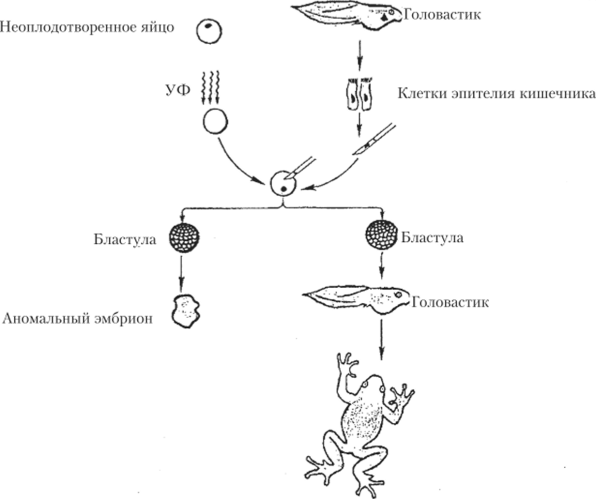
Рис. 10.1. Тотипотентность ядра соматической кпстк" Xenopus Iaevis.
Тотипотентность соматических клеток растений дает большие возможности для изучения дифференциального действия генов в онтогенезе. Так, сравнивая спектры изозимов, например пероксидазы, в недифференцированной каллусной ткани и в дифференцированных органах (корнях, листьях и т. д.) регенерантов, можно убедиться, что в каллусах образуется максимальный спектр изозимов пероксидазы, а в листьях или корнях целого растения спектр изозимов сужается. При дифференцировке происходит репрессия синтеза некоторых изозимов (рис. 10.2).
Дедифференцировка (каллус) сопровождается расширением изозимного спектра по сравнению со спектром дифференцированных органов, а вторичная дифференцировка вновь приводит к сужению спектра изозимов.
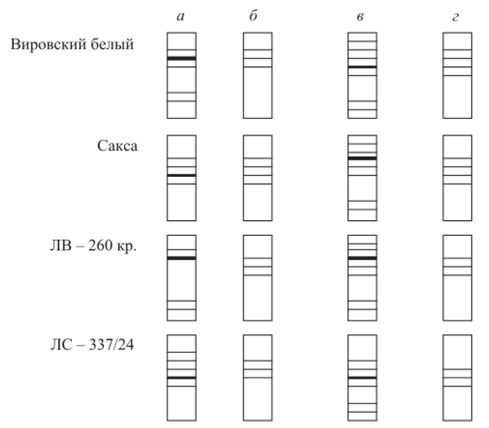
Рис. 10.2. Изозимные спектры пероксидазы редиса: а — семядоли;
6 — корни; в — каллус в культуре; г — корни, развившиеся в результате вторичной дифференцировки в каллусной культуре. Слева — наименования сортов и линии редиса Сравнение яйца или гаструлы с соматическими клетками животных показывает, что в первом случае синтезируется гораздо больше различных типов иРНК, чем во втором. Таким образом, встает вопрос об уровнях и механизмах обеспечения дифференциальной экспрессии генов.