Дыхание как саморегулируемый процесс

Альтернативное дыхание можно рассматривать как механизм поддержания баланса между углеводным обменом и скоростью транспорта электронов. Интенсивный транспорт электронов приводит к увеличению количества АТФ в матриксе и уменьшению количества АДФ. Это изменение соотношения АТФ и АДФ вызывает торможение электронного транспорта и накопление в матриксе митохондрии НАДН. Увеличение концентрации НАДН… Читать ещё >
Дыхание как саморегулируемый процесс (реферат, курсовая, диплом, контрольная)
Потребности клетки в АТФ, а также в промежуточных продуктах дыхания непрерывно меняются. Следовательно, растение должно иметь возможность регулировать дыхание, чтобы не тратить зря дыхательный субстрат, т. е. скорость его распада должна соответствовать потребностям клеток в АТФ и других продуктах дыхания.
Регуляция дыхания, как и других функций, осуществляется с помощью разных механизмов. Лучше других изучена регуляция активности ферментов. Если в клетке быстро идут процессы, для которых необходима энергия, то АТФ превращается в АДФ и неорганический фосфат. Увеличение концентрации фосфата в клетке активирует работу трех ферментов гликолиза: гексокиназы, фосфофруктокиназы и глицеральдегид- 3-фосфатдегидрогеназы, — и процесс пойдет быстрее. Однако большие количества АТФ тормозят гликолиз, дезактивируя фосфофруктокиназу. Одновременно происходит торможение пентозофосфатного окислительного цикла, так как увеличение количества фосфата снижает активность глюкозо-6-фосфатдегидрогеназы, транскетолазы и грансальдолазы. Увеличение количества фосфата является причиной усиления гликолиза в условиях гипоксии.
Увеличение количества АТФ тормозит цикл Кребса, так как снижается активность пируваткиназы и цитратсинтазы. Замедление цикла Кребса приводит к уменьшению количества АТФ, в результате ее ингибирующее действие снимается, и цикл ускоряется.
Работа дегидрогеназ цикла Кребса зависит от соотношения НАД+/НАДН в матриксе. Цитратсинтаза ингибируется НАДН и АТФ. Чем активнее цитратсинтаза, тем быстрее пойдут и другие реакции цикла, например дегидрирование веществ с восстановлением НАД. Однако увеличение количества НАДН вызовет ингибирование фермента, и цикл затормозится.
Следовательно, главную роль в управлении ЦТК играет соотношение НАДН/НАД+ и АТФ/АДФ в матриксе митохондрии. При высоком содержании АТФ и НАДН реакции цикла будут тормозиться.
Скорость гликолиза, цикла Кребса, а также расщепления жирных кислот, транспорта электронов зависит от соотношения АТФ и АДФ. Все эти реакции идут быстрее, когда отношение АТФ/АДФ уменьшается из-за усиленного расходования АТФ. АТФаза на внутренней мембране митохондрии работает быстрее, когда концентрация ее субстратов — АДФ и неорганического фосфата — увеличивается. Чем выше скорость фосфорилирования АДФ с образованием АТФ, тем больше протонов переходит в матрикс. В результате электрохимический градиент уменьшается, а его уменьшение, в свою очередь, приводит к ускорению транспорта электронов. Зависимость дыхательных процессов от соотношения количеств АТФ и АДФ была названа дыхательным контролем. При высоком уровне
АДФ и фосфата, который связан с быстрым расходом в клетке АТФ, ключевые реакции гликолиза и ЦТК активируются, а при накоплении АТФ тормозятся.
Механизм дыхательного контроля основан на том, что повышение отношения ЛТФ/АДФ замедляет синтез АТФ. Это соотношение определяется скоростью расхода АТФ. Скорость переноса электрона в цепи прямо зависит от работы АТФазы, т. е. от соотношения АТФ и АДФ в матриксе. При высоком соотношении АТФ и АДФ в матриксе АТФ аллостерически ингибирует активность цитохромоксидазы, что снижает активность всего комплекса IV.
Альтернативное дыхание можно рассматривать как механизм поддержания баланса между углеводным обменом и скоростью транспорта электронов. Интенсивный транспорт электронов приводит к увеличению количества АТФ в матриксе и уменьшению количества АДФ. Это изменение соотношения АТФ и АДФ вызывает торможение электронного транспорта и накопление в матриксе митохондрии НАДН. Увеличение концентрации НАДН блокирует цикл Кребса, в результате увеличивается концентрация пирувата, который активизирует альтернативную оксидазу, что приводит к быстрому окислению избытка НАДН. Таким образом, цикл Кребса возобновится, а его промежуточные вещества будут использоваться для других синтезов.
В регуляции дыхания участвуют и продукты реакции. Например, накопление ФЕП останавливает распад глюкозы, пируваткиназа ингибируется метаболитами цикла Кребса — цитратом и малатом, а изоцитратдегидрогеиаза, наоборот, активируется цитратом. Это примеры регуляции дыхания по принципу обратной связи. Обратной связью называют влияние более позднего члена цени взаимосвязанных реакций на более ранний член. Ценные и циклические процессы при помощи обратной связи могут регулироваться за счет продуктов, имеющихся в минимуме.
Современное представление о дыхании как о саморегулируемом процессе позволило объяснить знаменитый эффект Пастера: значительное уменьшение потребления глюкозы в присутствии кислорода воздуха объясняется тем, что при окислении НАДН дополнительно образуется АТФ. Поэтому в аэробных условиях клетке для удовлетворения своих энергетических потребностей достаточно меньшего количества глюкозы. Как клетка узнает, что ей можно расходовать меньше глюкозы? В присутствии кислорода усиливается образование АТФ и соответственно уменьшается концентрация неорганического фосфата, а последнее, как мы уже говорили, тормозит гликолиз.
Регуляция скорости дыхания в результате изменения концентрации метаболитов, получила название метаболической.
Дыхательные циклы открыты, т. е. отдельные вещества могут входить в цикл и выходить из него, влияя на скорость и химизм дыхания. В разных процессах — гликолиз и С3-цикл фотосинтеза, гликолиз и С4-цикл, пентозофосфатный окислительный цикл и С3-цикл — могут возникать одни и те же вещества (ФГК, ФГА, ФЕП, рибозо-5-фосфат, эритрозо-4-фосфат и др.). Благодаря существованию общих промежуточных веществ отдельные пути их превращения связываются в сетки реакций. Сетки реакций это средство для саморегулирования метаболизма. Многие ферменты конкурируют за один и тот же субстрат, поэтому в местах пересечения разных путей обмена веществ, т. е. там, где возникает общий продукт, появляется возможность переключения с одного пути обмена на другой. Замедление реакций одного цикла сопровождается ускорением реакций другого, что приводит к переключению с одного дыхательного пути на другой. Например, ФЕП может превращаться в ПВК во время гликолиза или карбоксилироваться с образованием оксалоацетата (С4-цикл, САМ-цикл):
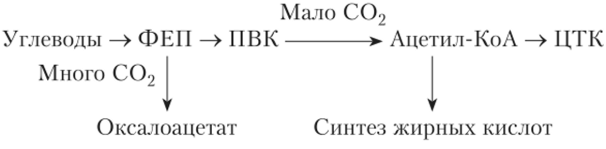
Следующий механизм регуляции дыхания связан с конформационными изменениями белков мембран митохондрий. В результате изменения формы белковых глобул происходит связывание или освобождение мембранами дыхательных ферментов, что влияет на их активность. Мембраны клетки, изменяя под влиянием различных раздражителей свою проницаемость, регулируют транспорт промежуточных веществ из одного компартмента в другой и их включение в соответствующий цикл. Увеличение проницаемости мембран митохондрий ускоряет поступление в эту органеллу из цитозоля продуктов гликолиза, поэтому цикл Кребса, а также образование АТФ пойдут быстрее. Увеличение проницаемости мембран во время хранения, например яблок, приводит к выделению дыхательных ферментов из митохондрий в цитозоль, что нарушает дыхание.
Однако основной механизм регуляции дыхания, как и других функций растительного организма, — это генетическая регуляция, например регуляция синтеза дыхательных ферментов (см. параграф 2.6).
Все эти механизмы действуют на клеточном уровне. Однако, как фотосинтез и водный обмен, дыхание регулируется и на других уровнях. Устьичные движения, скорость роста органа регулируют дыхание на органном уровне, а скорость поглощения и транспорта веществ — на уровне целого растения.
Дыхание — один из наиболее тонко регулируемых процессов.
Из сказанного в двух последних параграфах вытекает следующее.
- 1. Скорость дыхания относительно постоянна. Она поддерживается цикличностью дыхательного метаболизма, наличием регуляторных ферментов, а также стабилизирующей ролью наследственного аппарата.
- 2. Под действием раздражителей возникает нарушение стационарного состояния: период высокой активности сменяется угнетением процесса. Если раздражитель умеренной силы или раздражение непродолжительно, то устанавливается новый стационарный уровень интенсивности дыхания.
- 3. Раздражителями могут быть чередование температур, света и темноты, обезвоживание, механические раздражения, смена солевого состава питательного раствора, изменение газового состава атмосферы и другие факторы.
- 4. В экстремальных условиях температуры, влажности, под влиянием механических повреждений, вредных веществ или их концентраций, болезни, гипоксии скорость дыхания повышается — выделяется больше С02, но уменьшается физиологическая эффективность дыхания, нарушается синтез АТФ. Все эти неблагоприятные факторы вызывают одновременно изменение дыхательного коэффициента.
Контрольные вопросы
- 1. Как растение регулирует дыхание?
- 2. Какие механизмы регуляции дыхания известны?
- 3. Как регулируется активность дыхательных ферментов?
- 4. Что такое обратная связь?
- 5. Что такое дыхательный контроль?
- 6. Какое значение имеет цикличность дыхательных процессов?
- 7. На каких уровнях растение может регулировать дыхание?
Литература
Основная
- 1. Полевой, В. В. Физиология растений / В. В. Полевой. — М.: Высшая школа, 1989.
- 2. Скулачев, В. П. Кислород в живой клетке: добро и зло / В. П. Скулачев // Соровский образовательный журнал. — 1996. — № 3. — С. 4—10.
Дополнительная
- 1. Рахманкулова, 3. Ф. Дыхательные митохондриальные суперкомплексы у растений: структура и возможные функции / 3. Ф. Рахманкулова // Физиология растений. — 2014. - Т. 60. — С. 765−777.
- 2. Duncan, О. The outer mitochondrial membrane in higher plants / O. Duncan [et al. J // Trends in Plant Science. — 2013. - V. 18. — P. 207−217.
- 3. Schertl, P. Respiratory electron transfer pathways in plant mitochondria / P. Schertl, H.-P. Braun // Front. Plant Sci. — 2014. — doi: 10.3389/fpls.2014.163.