Вертикальное распределение зоопланктона термически стратифицированных озер Челябинской области

На рис. 8 показано вертикальное распределение кладоцер в разных водоемах. Диаграммы построены вышеописанным способом и не предназначены для сравнения численности представителей различных видов на каком-либо горизонте. Типичное распределение кладоцер характеризуется абсолютным максимумом в приповерхностном слое и локальным максимумом в гиполимнионе или в нижней части металимниона, но не… Читать ещё >
Вертикальное распределение зоопланктона термически стратифицированных озер Челябинской области (реферат, курсовая, диплом, контрольная)
ВЕРТИКАЛЬНОЕ РАСПРЕДЕЛЕНИЕ ЗООПЛАНКТОНА ТЕРМИЧЕСКИ СТРАТИФИЦИРОВАННЫХ ОЗЕР ЧЕЛЯБИНСКОЙ ОБЛАСТИ
В настоящее время совершенно очевидно, что бесконтрольная деятельность человека ведет к крайне негативным последствиям, связанным со снижением несущей способности экосистем. В результате за сравнительно небольшой промежуток времени (несколько лет или десятилетий) происходит трансформация, сопоставимая с естественными изменениями, охватывающими целые геологические эпохи. Для того чтобы выявить антропогенные изменения на фоне естественных флуктуаций, необходима достаточно подробная и разносторонняя информация.
Одной из важнейших особенностей водных экосистем является их вертикальная неоднородность, связанная с поступлением света и биогенов, аэрацией, перемещением донных осадков под действием силы тяжести. Вместе с тем, изучение зоопланктона — сообщества, от состояния которого зависят потребительское качество воды и рыбопродуктивность водоема, — традиционно строится на анализе средневзвешенных проб или проб, полученных в результате тотального облова столба воды планктонными сетями. Материалы, основанные на анализе результатов послойного лова зоопланктона, встречаются довольно редко. Относительно озер Челябинской области таких сведений в открытой печати обнаружить не удалось.
Настоящая работа посвящена изучению вертикального распределения зоопланктона озер Увильды, Большое Миассово, Малый Теренкуль в период летней стратификации.
Озера, выбранные для исследования, имеют тектоническое происхождение и расположены в пределах района Восточных предгорий Южного Урала [1]. Эти озера в различной степени подвержены антропогенному воздействию.
Озеро Увильды — самый крупный естественный водоем на Южном Урале, его площадь составляет 68,0 км², объем 1014 млн м3, средняя глубина — 14 м при максимальной 38 м. Для этого озера характерны глубокая котловина с крутыми стенками, разделенная подводными возвышениями на три части, слабо развитая литораль, изрезанная береговая линия, сложный рельеф дна с многочисленными скалистыми грядами [1]. Несмотря на то что озеро с середины XX в. интенсивно используется в рекреационных целях, его побережье и водосборная площадь в значительной мере застроены, оно сохраняет признаки олиготрофного водоема [4]. В период летней стратификации практически вся его толща насыщена кислородом до предельного уровня.
Озеро Б. Миассово представляет собой западный плес единого озера Миассово, его площадь 11,4 км², объем водной массы 129 млн м3. Максимальная глубина озера составляет 25 м, средняя — 11 м. Наличие большого количества заливов и разнообразное строение берегов обусловили высокую изрезанность береговой линии. Прибрежная зона развита слабо, сразу же от берега глубины резко увеличиваются. Дно неровное, с выходами горных пород в виде подводных островов, скал, гольцов, разделенных сапропелевыми илами [5]. Озеро Б. Миассово является центральным звеном Кисегач-Миассовской озерной системы, крупнейшим водоемом Ильменского заповедника. Это озеро можно рассматривать как эталон озерной экосистемы, в малой степени подверженной влиянию человеческой деятельности. В период летней стагнации вся толща эпилимниона равномерно насыщена кислородом, относительные показатели концентрации которого колеблютсяв пределах, близких к насыщению (98−110%). Содержание кислорода летом в придонных слоях составляет 89−99% насыщения. В отличие от озера Увильды, озеро Б. Миассово является мезотрофным, хотя и сохраняет некоторые черты олиготрофии [5].
Озеро Малый Теренкуль — небольшой водоем площадью 0,98 км², объемом около 9 млн м3; максимальная глубина — 19 м, средняя — 9 м. Донный рельеф однообразен и представляет собой котловину с единым центром. Сочетание небольшой площади озера со значительной глубиной, а также закрытые, поросшие лесом берега способствуют формированию в летнее время четко выраженной термической стратификации. Начиная с 1926 г. в озеро поступают хозбытовые стоки со стороны рекреационных объектов курортной зоны [2]. На момент отбора проб прозрачность по диску Секки была не больше 1 м. Начиная с глубины 5 м содержание кислорода не превышало 0,5 мг/л, отмечено появление сероводорода. По современному состоянию данное озеро может быть отнесено к переходному гипер-политрофному типу [6].
Сбор исходных материалов осуществлялся в следующем порядке: 9.07.2007 были отобраны пробы в зоне максимальных глубин на озере Увильды (станция 2, рис. 1), 8.07.2008 на данном водоеме удалось отобрать пробы на пяти станциях (1−5). Работы на озерах М. Теренкуль и Б. Миассово проводились и 27.07.2010 соответственно, положение станций показано на рис. 1.
Для лова зоопланктона использовалась стандартная количественная сеть Джеди (диаметр верхнего кольца — 18 см, нижнего — 24 см, фильтрующий материал — газ-сито № 64). Облавливался весь столб воды послойно. При отборе проб на озере Увильды выделялись горизонты с интервалом 5 м, на озерах М. Теренкуль и Б. Миассово — 3 м. Видовой состав определялся с использованием соответствующей литературы. Для оценки численности и биомассы зоопланктона применялся стандартный счетный метод [3].
В ходе исследования вертикального распределения зоопланктона озера Увильды обнаружено 25 видов гидробионтов: 11 видов — представители отряда Cladocera, класс Crustacea (Ракообразные); 6 видов — представители отряда Copepoda, класс Crustacea; 8 видов относятся к типу Rotifera (Коловратки). Видовой состав, численность, биомасса и приуроченность каждого вида к выделенным горизонтам приведены в табл. 1 и 2. Из таблиц следует, что видовой состав по материалам, собранным в разные годы, не совпадает. Так, в 2007 г. отмечено больше видов ветвистоусых и веслоногих ракообразных, тогда как в 2008 г. возросло число видов коловраток. Подобные изменения могут быть связаны с динамикой численности отдельных видов, с особенностями их жизненных циклов, а также с погодными условиями. Средневзвешенные значения численности составили 36,0 тыс. экз./м3 и 21,0 тыс. экз./м3 для сезонов 2007 и 2008 г. соответственно, биомассы — 0,50 г/м3 и 0,57 г/м3.
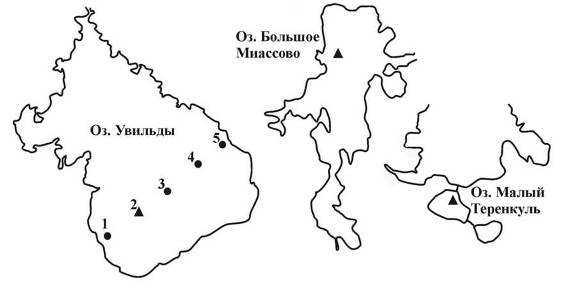
Рис. 1. Расположение станций отбора проб; треугольником отмечены станции в зоне максимальных глубин
Таблица 1.
Видовой состав и количественные характеристики зоопланктона на разных горизонтах в толще воды озера Увильды по результатам наблюдений 9.07.2007.
Виды. | Горизонт, м. | ||||
0−5. | 5−10. | 10−15. | 15−20. | 20−30. | |
Отр. Cladocera, кл. Crustacea. | |||||
Daphnia longispina. | |||||
(O. F. Muller, 1785). | 2028,3. | 268,7. | 23,5. | 58,3. | 1,4. |
Ceriodaphnia quadrangula. | |||||
(Muller, 1785). | 17,1. | 3,0. | 1,7. | ||
Bosmina longirostris. | |||||
(Muller, 1785). | 12,8. | 78,6. | 19,2. | 33,5. | 8,2. |
Chydorus sphaericus. | |||||
(Muller, 1785). | 1,0. | 0,5. | 1,3. | 1,2. | 1,3. |
Diaphanosoma brachiurum. | 49,2. | ||||
(Levin, 1848). | 56,2. | 18,2. | 0,2. | 3,3. | 0,5. |
Leptodora kindtii. | |||||
(Focke, 1844). | 39,0. | 3,1. | 0,2. | 3,5. | |
Polyphemus pediculus. | |||||
(Linneus, 1778). | 51,6. | 7,1. | 0,7. | 36,0. | |
Bythotrephes longimanus. | |||||
(Leydig, 1860). | 0,4. | ||||
Отр. Copepoda, кл. Crustacea. | |||||
Mesocyclops leuckarti. | |||||
(Claus, 1857). | 0,9. | 2,2. | |||
Thermocyclops oithonoides. | |||||
(Sars, 1863). | 132,9. | 111,4. | 3,9. | 5,9. | |
Копеподиты циклопид. | |||||
младших стадий. | 13,7. | 167,8. | 51,5. | 59,5. | 18,1. |
Eudiaptomus graciloides. | |||||
(Lilljeborg, 1889). | 55,2. | 14,2. | 2,4. | 8,7. | |
Nauplii. | 13,7. | 1,8. | 1,4. | 2,5. | 1,6. |
Тип Rotifera. | |||||
Bipalpus hudsoni. | |||||
(Imhof, 1891). | 5,5. | 18,3. | 15,6. | 5,0. |
- * В числителе — количество, экз./м3; в знаменателе — биомасса, мг/м3.
- ** Помимо перечисленных видов в пробах отмечены единичные экземпляры Sida crystalline (Muller, 1776), Kellicottia longispina (Kellicott, 1879), Keratella cochlearis (Gosse, 1851), Filinia longiseta (Ehrenberg, 1834), численность которых не определялась.
Таблица 2.
Видовой состав и количественные характеристики зоопланктона на разных горизонтах в толще воды озера Увильды по результатам наблюдений 8.07.2008.
Виды. | Горизонт, м. | |||||
0−5. | 5−10. | 10−15. | 15−20. | 20−25. | 25−30. | |
Отр. Cladocera, кл. Crustacea. | ||||||
Daphnia longispina. | ||||||
(O. F. Muller, 1785). | 21,6. | 11,2. | 1,1. | 2,1. | 1,5. | 1,0. |
Daphnia ^cullata. | ||||||
(Sars, 1862). | 1964,9. | 383,5. | 18,7. | 17,5. | 45,1. | 5,3. |
Ceriodaphnia quadrangula. | 30.0. | |||||
(Muller, 1785). | 4,2. | 4,4. | 0,4. | 0,2. | ||
Daphnia cristata. | ||||||
(Sars, 1862). | 16,1. | 4,2. | 1,9. | 0,7. | 1,6. | 1,6. |
Bosmina longirostris. | ||||||
(Muller, 1785). | 21,6. | 68,3. | 94,8. | 65,5. | 173,4. | 109,0. |
Chydorus sphaericus. | ||||||
(Muller, 1785). | 0,7. | 0,2. | 0,3. | 0,1. | ||
Diaphanosoma brachiurum. | ||||||
(Levin, 1848). | 56,2. | 4,4. | 0,2. | 1,0. | 0,2. | |
Leptodora kindtii. | ||||||
(Focke, 1844). | 39,2. | 13,8. | 8,9. | |||
Polyphemus pediculus. | ||||||
(Linneus, 1778). | 16,64. | 2,65. | ||||
Отр. Copepoda, кл. | . Crustacea. | |||||
Сyclops vicinus. | ||||||
(Uljanin, 1875). | 1,4. | 7,4. | 5,9. | 3,9. | 5,8. | 6,4. |
Mesocyclops leuckarti. | ||||||
(Claus, 1857). | 7,3. | 7,3. | 0,5. | 0,2. | 0,8. | 0,4. |
Thermocyclops oithonoides. | ||||||
(Sars, 1863). | 104,9. | 75,0. | 2,0. | 1,2. | 1,6. | 0,7. |
Копеподиты циклопид. | ||||||
младших стадии. | 1,7. | 2,9. | 1,1. | 1,5. | 2,0. | 1,2. |
Eudiaptomus graciloides. | ||||||
(Lilljeborg, 1889). | 30,0. | 1,3. | 0,7. | 0,3. | 1,0. | 0,1. |
Nauplii. |
|
|
|
|
|
|
Тип Rotifera. | ||||||
Kellicottia longispina. | ||||||
(Kellicott, 1879). | 0,1. | *. | *. | *. | *. | *. |
Keratella cochlearis. | ||||||
(Gosse, 1851). | *. | |||||
Keratella quadrata. | ||||||
(O.F. Muller, 1786). | *. | *. | 0,1. | 0,2. | 1,1. | 0,1. |
Filinia longiseta. | ||||||
(Ehrenberg, 1834). | 0,1. | 0,8. | 0,6. | 0,1. | ||
Asplanchna priodonta. | ||||||
(Gosse, 1850). | 10,2. | 0,8. | ||||
Bipalpus hudsoni. | ||||||
(Imhof, 1891). | 0,3. | 2,4. | 6,8. | 1,5. | 1,6. | 0,6. |
* В числителе — количество, экз./м3; в знаменателе — биомасса, мг/м3; символом «*» обозначены показатели биомассы менее 0,05 мг/м3.
Характеристики видового состава, численности и биомассы зоопланктона озера Б. Миассово приведены в табл. 3. Всего обнаружено 23 вида гидробионтов: 8 видов Cladocera, 5 видов Copepoda и 10 видов Rotifera. Видовой состав гидробионтов озер Увильды и Б. Миассово аналогичен и в целом характерен для пресных озер Южного Урала. Различия касаются некоторых деталей. Так, массовое присутствие представителей рода Cyclops в середине лета, к тому же в наиболее прогретом слое, — явление необычное. Представители данного рода, как правило, дицикличны и наиболее многочисленны весной и поздней осенью. Также примечательна высокая численность Chydorus sphaericus — эвритермного вида, постоянно присутствующего в составе зоопланктона, но не относящегося к числу доминирующих. Усредненные для всего столба воды значения численности и биомассы составили 39,6 тыс. экз./м3 и 1,05 г/м3 соответственно. Таблица 3.
вертикальный распределение зоопланктона озеро Видовой состав и количественные характеристики зоопланктона на разных горизонтах в толще воды озера Б. Миассово.
Го. | эизонт, м. | ||||||
Виды. | 0−3. | 3−6. | 6−9. | 9−12. | 12−15. | 15−18. | 18−21. |
Отр. Cladocera,. | кл. Crustacea. | ||||||
Daphnia longispina. | |||||||
(O. F. Muller, 1785). | 21,9. | 15,0. | 12,6. | 0,8. | |||
Daphnia cucullata. | |||||||
(Sars, 1862). | 2623,5. | 1261,9. | 653,9. | 41,2. | 17,1. | 27,9. | |
Ceriodaphnia quadrangula. | |||||||
(Muller, 1785). | 0,1. | ||||||
Daphnia cristata. | |||||||
(Sars, 1862). | 0,2. | 16,2. | 0,2. | 0,9. | 0,3. | ||
Bosmina kessleri. | |||||||
(Muller, 1785). | 49,5. | 0,7. | 66,0. | 2,7. | 4,9. | 2,1. | 3,3. |
Chydorus sphaericus. | |||||||
(Muller, 1785). | 228,8. | 18,1. | 29,5. | 3,2. | 6,0. | 1,8. | 2,8. |
Diaphanosoma brachiurum. | |||||||
(Levin, 1848). | 265,7. | 22,6. | 4,3. | 0,5. | 3,2. | 0,6. | 0,8. |
Leptodora kindtii. | |||||||
(Focke, 1844). | 94,9. | 44,7. | 74,0. | 5,7. | 0,9. | ||
Bythotrephes longimanus. | |||||||
(Leydig, 1860). | 132,7. | 9,4. | 19,0. | 2,7. | 6,5. | ||
Отр. Copepoda,. | кл. Crustacea. | ||||||
Cyclops sp. | |||||||
772,5. | 29,6. | 1,3. | 1,4. | 0,8. | |||
Mesocyclops leuckarti. | |||||||
(Claus, 1857). | 80,2. | 70,7. | 10,9. | 8,4. | 1,3. | 3,9. | |
Thermocyclops oithonoides. | |||||||
(Sars, 1863). | 150,9. | 103,7. | 6,3. | 8,0. | 3,5. | 3,5. | |
Копеподиты циклопид. | |||||||
младших стадий. | 93,1. | 22,3. | 59,4. | 4,1. | 2,5. | 2,0. | 1,7. |
Eudiaptomus graciloides. | |||||||
(Lilljeborg, 1889). | 30,0. | 9,9. | 34,3. | 2,0. | 5,9. | 2,5. | 3,8. |
Nauplii. | |||||||
5,0. | 3,5. | 2,5. | 0,2. | 0,1. | 0,1. | 0,1. | |
Тип Rotifera. | |||||||
Kellicottia longispina. | |||||||
(Kellicott, 1879). | *. | 0,1. | *. | *. | *. | ||
Keratella cochlearis. | |||||||
(Gosse, 1851). | *. | *. | *. | *. | *. | *. | |
Keratella quadrata. | |||||||
(O. F. Muller, 1786). | *. | *. | *. | *. | *. | ||
Filinia longiseta. | |||||||
(Ehrenberg, 1834). | 0,2. | 0,1. | *. | ||||
Asplanchna priodonta. | |||||||
(Gosse, 1850). | 1,0. | 0,8. | |||||
Euchlanis dilitata. | |||||||
(Ehrenberg, 1832). | 0,9. | 0,8. | *. | *. | *. | *. | *. |
Polyartra sp. |
|
| ; |
| ; |
| ; |
Trichocerca sp. | ; | ; |
|
|
|
|
|
* В числителе — количество, экз./м3; в знаменателе — биомасса, мг/м3; символом «*» обозначены показатели биомассы менее 0,05 мг/м3.
Видовой состав и количественные характеристики зоопланктона на разных горизонтах в толще воды озера М. Теренкуль.
Виды. | Горизонт, м. | |||||
0−3. | 3−6. | 6−9. | 9−12. | 12−15. | 15−18. | |
Отр. Cladocera, кл. | Crustacea. | |||||
Daphnia longispina. | ||||||
(O. F. Muller, 1785). | 2970,5. | 266,0. | 89,1. | 85,4. | 32,02. | 38,2. |
Bosmina longirostris. | ||||||
(Muller, 1785). | 0,9. | 0,1. | ||||
Leptodora kindtii. | ||||||
(Focke, 1844). | 3,8. | 25,9. | 0,5. | 1,0. | ||
Bythotrephes longimanus. | ||||||
(Leydig, 1860). | 25,0. | |||||
Отр. Copepoda, кл. | Crustacea. | |||||
Mesocyclops leuckarti. | ||||||
(Claus, 1857). | 2,1. | 7,0. | 0,2. | 0,1. | ||
Thermocyclops oithonoides. | ||||||
(Sars, 1863). | 218,6. | 106,7. | 3,5. | 10,0. | 5,0. | 4,7. |
Копеподиты циклопид. | ||||||
младших стадии. | 12,7. | 6,8. | 0,3. | 0,3. | *. | 0,3. |
Eudiaptomus graciloides. | ||||||
(Lilljeborg, 1889). | 1,4. | 1,6. | ||||
Nauplii. |
|
|
|
|
|
|
Тип Rotifera. | ||||||
Brachionus diversicornis. | ||||||
(Daday, 1883). | 8,9. | 1,2. | 2,0. | 1,1. | 0,2. | 0,6. |
Keratella cochlearis. | ||||||
(Gosse, 1851). | 0,1. | 0,1. | 0,1. | 0,1. | 0,1. | 0,1. |
Keratella quadrata. | ||||||
(O. F. Muller, 1786). | 2,52. | 0,2. | 0,6. | 0,1. | 0,1. | 0,1. |
Euchlanis dilitata. | ||||||
(Ehrenberg, 1832). | *. | *. | *. | |||
Lecane luna. | ||||||
(O. F. Muller, 1776). | 0,1. | *. | *. | *. | *. | *. |
* В числителе — количество, экз./м3; в знаменателе — биомасса, мг/м3; символом «*» обозначены показатели биомассы менее 0,05 мг/м3.
В ходе исследования вертикального распределения зоопланктона озера М. Теренкуль обнаружено 13 видов гидробионтов: 4 — Cladocera,.
— Copepoda, и 4 — Rotifera. Видовой состав, численность, биомасса и приуроченность каждого вида к выделенным горизонтам приведены в табл. 4. Из таблицы следует, что видовой состав зоопланктона достаточно беден, причем только в отношении одного вида — Brachionus diversicornis (Daday, 1883) можно сказать, что он является высокосапробным, т. е. характерным для вод, загрязненных органикой. Представителей остальных видов мы с полным основанием причисляем к эврибионтным.
Главной отличительной чертой зоопланктона озера М. Теренкуль следует считать отсутствие целого ряда характерных для данного класса озер видов: Daphnia cristata (Sars, 1862), Daphnia cucullata (Sars, 1862), Diaphanosoma brachiurum (Levin, 1848), Ceriodaphnia quadrangula (Muller, 1785). Кроме того, обращает на себя внимание, что такие массовые виды, как Eudiaptomus graciloides (Lilljeborg, 1888) и Bosmina longirostris (Muller, 1785), представлены единичными экземплярами.
Усредненная численность зоопланктона составила 25,8 тыс. экз./м3, биомасса — 0,66 г/м3. Необходимо отметить, что, несмотря на значительно более высокий трофический статус озера М. Теренкуль, развитие зоопланктона, оцениваемое по показателям обилия, приблизительно соответствует таковому для озера Увильды и явно Таблица 4уступает озеру Б. Миассово. Наблюдаемые различия можно объяснить следующим образом. В озере Увильды, сохраняющем черты олиготрофии, фактором, лимитирующим развитие зоопланктона, выступает обеспеченность пищей, а в гипертрофном озере М. Теренкуль — токсичное воздействие выделений сине-зеленых водорослей и продуктов неполного окисления органики. В озере Б. Миассово действие этих факторов менее выражено.
Параллельно с ловом зоопланктона проводилось измерение температуры. Вертикальное распределение температур представлено на рис. 2.
Приведенные графики позволяют проследить положение металимниона. В толще воды озера М. Теренкуль он располагается в слое от 3 до м. Для озера Б. Миассово характерен более размытый металимнион, охватывающий интервал 3−10 м. По данным наблюдений 9.07.2007, металимнион в озере Увильды располагался в слое 10−20 м. По всей видимости, в период, предшествующий проведению съемки 8.07.2008, имело место значительное похолодание. В результате слой скачка температур оказался размытым (рис. 2). На это же обстоятельство указывает положение изотерм 15° и 13° на рис. 3, из которого следует, что прогрев охватывает более глубокие слои в центральной части котловины, чем на ее периферии.
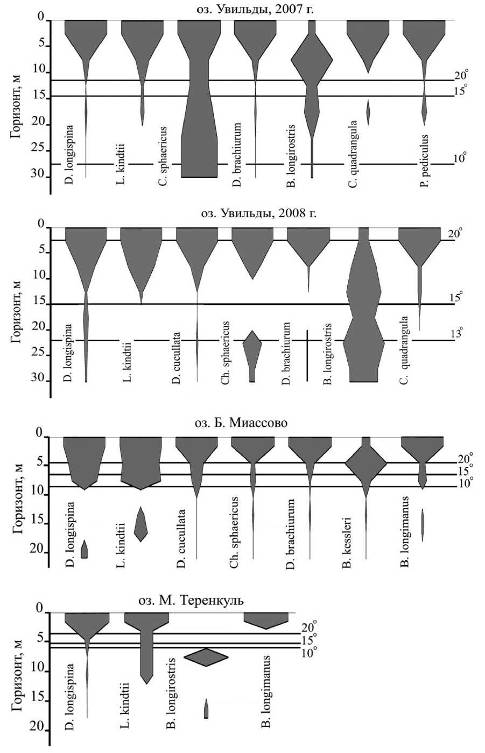
На рис. 3 и 4 показано вертикальное распре — деление зоопланктона во всех исследованных водоемах. Приведенные диаграммы построены следующим образом. После того как для каждой станции были определены значения численности гидробионтов в различных горизонтах, выбирали максимальное значение и принимали его равным единице, численность на других горизонтах этой же станции выражали в долях единицы. Каждое значение соотносилось со шкалой глубин. Считали, что численность в данном горизонте соответствует на графике середине интервала. Например, если значение численности гидробионтов получено в результате облова слоя воды от 10 до 5 м, то на графике данное значение будет соответствовать глубине 7,5 м. При этом необходимо отметить, что приведенные диаграммы позволяют анализировать особенности вертикального распределения гидробионтов, но не предназначены для сравнения их численности на разных станциях.
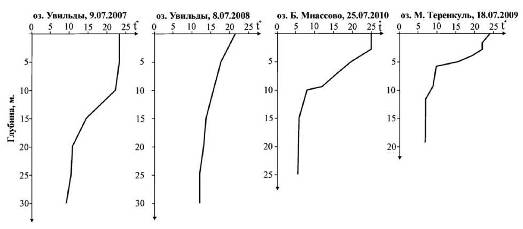
Рис. 2. Вертикальное распределение температур
Рис. 3. Вертикальное распределение планктонных организмов озера Увильды по результатам съемки 8.07.2008, пояснения в тексте
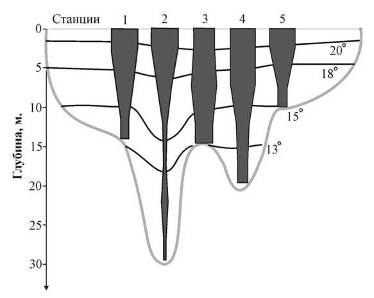
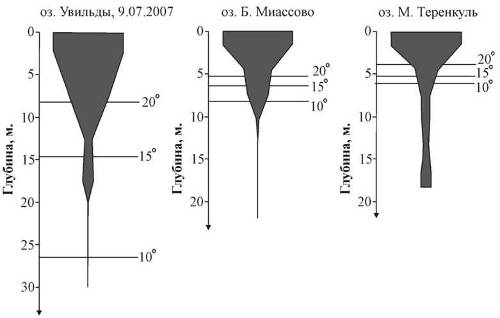
Рис. 4. Вертикальное распределение планктонных организмов по результатам послойного облова столба воды в самой глубокой части акватории, пояснения в тексте
Из рисунков следует, что общей тенденцией является концентрация гидробионтов в верхних, наиболее прогретых и обеспеченных пищей слоях. На равномерность распределения оказывает влияние также максимальная глубина в месте отбора проб. На рис. 3 видно, что наиболее неоднородно зоопланктон распределен в зоне, где глубины достигают 30 м.
Кроме того, на распределение влияют положение металимниона и величина перепада температур в его пределах. По всей видимости, наличие резко выраженной термической стратификации способствует концентрации гидробионтов в приповерхностном слое. Это отчетливо прослеживается на примере озер близкого трофического статуса Увильды и Б. Миассово. Что касается озера М. Теренкуль, то там картина оказывается несколько смазанной вследствие концентрации коловраток в аноксичном гиполимнионе.
Несомненный интерес представляет анализ соотношения численности основных групп зоопланктона в разных слоях водной толщи. На рис. 5 показано, как меняется доля ветвистоусых ракообразных (Cladocera) по мере увеличения глубины. Считается, что кладоцеры чувствительны к содержанию кислорода. Поэтому вполне закономерно, что в озере Увильды, где вся толща насыщена кислородом, их доля составляет от 63,5 до 86,6% и не уменьшается с глубиной.
Доля кладоцер в оз. Б. Миассово составляет от 28,8 до 59,7% в разных горизонтах и несколько снижается в глубоких слоях.
Принципиально иная ситуация складывается в озере М. Теренкуль: наибольшая доля кладоцер — 42,8% — приходится на самый верхний, насыщенный кислородом слой. По мере увеличения глубины их относительная численность падает, достигая 5,8% в придонном слое. Возникает вопрос: как вообще они могут существовать в бескислородном гиполимнионе? Сам факт проникновения кладоцер в бескислородный гиполимнион отражен в гидробиологической литературе [8]. При этом нам не удалось обнаружить описания экспериментов, которые подтверждали бы способность ветвистоусых к анаэробиозу.
В отличие от ветвистоусых коловратки в середине лета не являются доминирующей группой зоопланктона в эпилимнионе (рис. 6). Тем не менее, необходимо отметить весьма существенное увеличение их доли у нижней границы металимниона. Явное преобладание по численности коловраток в гиполимнионе озера М. Теренкуль может быть отнесено исключительно на счет загрязнения.
Напротив, изменение доли копепод в общей численности зоопланктона по мере увеличения глубины в относительно чистом озере Увильды и загрязненном озере М. Теренкуль практически не отличается (рис. 7). Это связано с тем, что основу копеподного комплекса обоих озер составляют одни и те же компоненты: T. oithonoides, M. leuckarti, копеподиты циклопид младших стадий, а также науплиусы. Эта же тенденция имеет место для копепод озера Б. Миассово, но присутствие в значительных количествах представителей р. Cyclops оказывает заметное влияние, и доля данной группы становится приблизительно одинаковой во всем столбе воды.
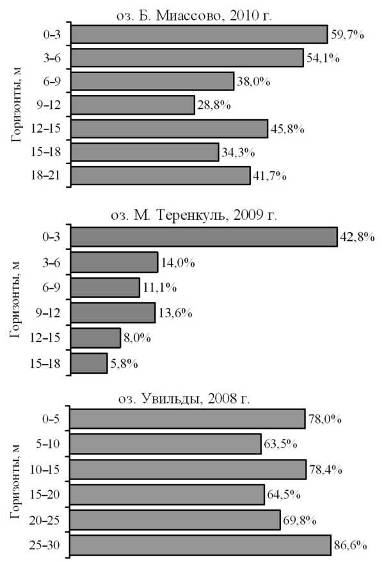
Рис. 5. Доля кладоцер в общей численности зоопланктона на разных горизонтах
На рис. 8 показано вертикальное распределение кладоцер в разных водоемах. Диаграммы построены вышеописанным способом и не предназначены для сравнения численности представителей различных видов на каком-либо горизонте. Типичное распределение кладоцер характеризуется абсолютным максимумом в приповерхностном слое и локальным максимумом в гиполимнионе или в нижней части металимниона, но не в придонном слое. Причем такой характер распределения присущ как круглогодичным видам (D. longispina, C. sphaericus), так и теплолюбивым летним видам (D. cucullata, D. brachiurum, С quadrangula, P. pediculus). Возможны некоторые отклонения: например, представители B. longimanus в озере Б. Миассово распределены вполне типично, а в озере М. Теренкуль нижний локальный максимум не обнаруживается. Это можно отнести на счет оксифильности данного вида, но не исключено, что его представители просто редко встречаются ниже термоклина.
Особо следует отметить характер распределения B. Longirosris. Представители данного вида совершают вертикальные суточные миграции, поэтому их максимальная плотность в дневное время оказывается на некоторой глубине, а в ночное время они поднимаются ближе к поверхности [7]. Примечательно, что максимум численности данного вида в озере Увильды приходится на горизонт от 5 до 10 м, а в озере М. Теренкуль — от 6 до 9 м, с той разницей, что в первом случае это зона эпилимниона, а во втором — гиполимниона.
Имееется также два максимума плотности представителей более теплолюбивого вида kessleri в озере Б. Миассово: один в пределах горизонта от 3 до 6 м (в металимнионе, у верхней границы), другой, слабовыраженный, — от 12 (в гиполимнионе).
На рис. 9 показано вертикальное распределение копепод. В типичном случае копеподы распределены аналогично кладоцерам — два максимума численности, выше и ниже металимниона. Причем круглогодично присутствующие в составе планктона представители эвритермных видов — E. graciloides, C. vicinus, копеподиты циклопид, достигающие половой зрелости в осеннее время, имеют тенденцию к более равномерному распределению, чем представители типично летнего вида — T. oithonoides. Хотя и для него обнаруживается слабо выраженный максимум ниже металимниона. Достаточно часто верхний максимум численности расположен в пределах металимниона. Это отмечено для T. oithonoides, M. leuckarti, а также копеподитов циклопид младших стадий.
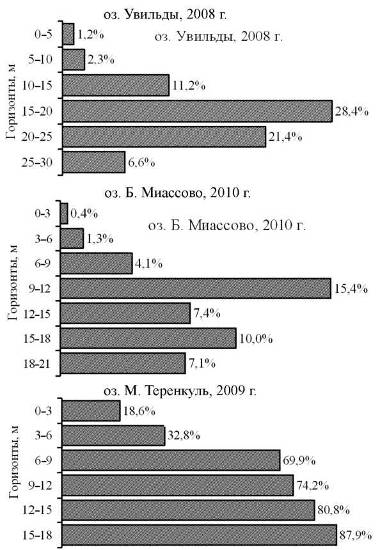
Рис. 6. Доля коловраток в общей численности зоопланктона на разных горизонтах
Распределение коловраток отличается большим разнообразием (рис. 10). В сезонном цикле массовое развитие коловраток происходит, как правило, в конце апреля — начале мая, когда с талыми водами поступает аллохтонная органика, или поздней осенью, после отмирания значительной части водорослей и макрофитов. В том и в другом случае коловратки наиболее многочисленны в холодных водах, поэтому их «привязка» к гиполимниону и нижней части металимниона вполне ожидаема. В действительности наблюдается сложная картина. Один и тот же вид может быть распределен по-разному, хотя некоторая закономерность все же прослеживается.
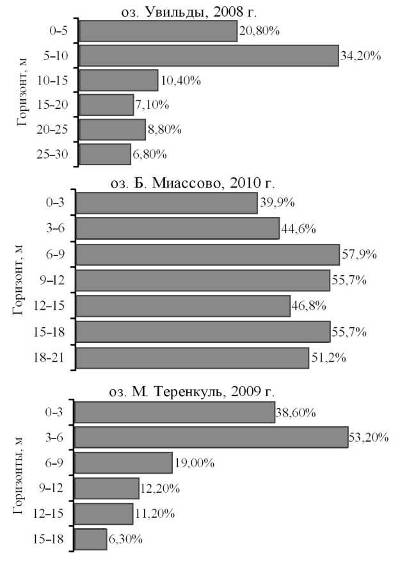
Рис. 7. Доля копепод в общей численности зоопланктона на разных горизонтах
K. longispina, по материалам изучения озера Увильды в 2008 г., имеет два максимума численности: в приповерхностном слое и в интервале глубин от 20 до 25 м. Этот же вид в озере Б. Миассово имеет только один максимум — в интервале 6−9 м. Глубинные слои, где наблюдается скопление коловраток данного вида, имеют сходную температуру — около 13 °C. Аналогично, F. longiseta предпочитает слои с температурой 13−15 °С. Максимум численности B. hudsoni приходится н…