Возрастные изменения белой пульпы селезенки у индеек в постнатальном онтогенезе

У 180-дневного возраста, скопления клеток в пульпе органа клетки располагаются наиболее рыхло, как в белой, так и в красной. Но, белая пульпа селезенки представлена функционально активными лимфоидными структурами, т. е. Ти В-зависимыми зонами. По сравнений со 150-дневными индейками, где отмечается незначительное уменьшение площади белой пульпы. Ретикулиновая сеть Ти В-зависимых зон разрыхлена… Читать ещё >
Возрастные изменения белой пульпы селезенки у индеек в постнатальном онтогенезе (реферат, курсовая, диплом, контрольная)
Возрастные изменения белой пульпы селезенки у индеек в постнатальном онтогенезе
С помощью гистоморфологических методов исследования изучены возрастные изменения белой пульпы селезенки у здоровых без породных индеек в постнатальном онтогенезе. Отмечены различные возрастные морфофункциональные состояния белой пульпы селезенки. При этом установлены сроки формирования и возрастные соотношение Ти Взависимых зон, выявлена максимальная морфофункциональная активность и возрастная физиологическая инволюция Ти Взависимых зон белой пульпы селезенки.
В отличие от млекопитающих, селезенка у птиц не выполняет функцию депо крови, и, начиная с момента появления цыпленка на свет, в ней происходят разрушение эритроцитов и образование лимфоцитов. Поэтому, в постнатальный период, селезенка у птиц является только органом лимфопоэза, и обеспечивает защиту организма от генетически чужеродных клеток, веществ, а также участвует в иммунных реакциях гуморального типа [1, 2].
Как и у других животных, в селезенке птиц различают белую и красную пульпы. Белая пульпа селезенки — в основном заселена соответствующими дифференцированными лимфоцитами. Участки белой пульпы включают в себя артериальные сосуды, окруженные преимущественно малыми лимфоцитами (Т — зависимая лимфатическая ткань), они не имеют четких границ, определенной конфигурации и величины. Кроме этого, в белой пульпе селезенки птиц есть лимфоидные фолликулы, сгруппированные лимфоидными клетками, они окружены четко выраженной соединительной тканью. Эти лимфоидные фолликулы относятся к В — зависимой лимфоидной ткани [3, 4, 5].
Красная, как и белая пульпа не имеет резких границ. Лимфоидные фолликулы сильно варьируют в зависимости от возраста и имеют слабо выраженные герминативные центры. Кроме этого, отличительная особенность белой пульпы селезенки индеек это то, что В-зависимая зона формируется позже по сравнению Тзависимой зоной и раньше подвергается возрастной физиологической инволюции [4,5].
Имеются отечественные научные работы, которые посвящены глубокому исследованию в норме морфофизиологии белой пульпы, ее Т — и Взависимых зон у разных видов животных и кур. Кроме этого, результаты исследований многих отечественных и зарубежных работ включают, что соотношение между белой и красной пульпой зависит от кровенаполнения селезенки и функционального состояния ее лимфоидной ткани, а также обусловлено видовыми особенностями [5, 6, 7,8].
Однако, многие вопросы касающиеся селезенки индеек изучены недостаточно. Поиск данных литературы по изучению онтогенеза селезенки у индеек не дал положительных результатов. Встречаются лишь отдельные сведения о морфологии селезенки кур и перепелов. По этому, суть данной научной работы — проследить изменения белой пульпы (Ти В — зон) селезенки индеек в постнатальном онтогенезе в разные периоды развития [5, 9, 10]. селезенка лимфопоэз птица Материалы и методы исследований Работа проводилась в период с 2005;2012гг. на кафедре «Ветеринарно-санитарная экспертиза, гистология и патология» Кыргызского Национального аграрного университета им. К. И. Скрябина, г. Бишкек. Объектом для исследования служили клинически здоровые цыплята-индейки (безпородные) двенадцати возрастных групп (однодневные, 7-, 14-, 21-, 30-, 60-, 90-, 120-, 150-, 180-, 270- и 420-дневные). Из каждой возрастной группы исследовалось по пять голов птицы. Материалом для исследования служила селезенка. Для выполнения поставленных задач использовались гистологические методы. При выполнении гистологических исследований пользовались руководством Г. А. Меркулова [11].
Результаты исследований
В процессе роста селезенки происходили следующие морфофункциональные изменения:
- · к моменту вылупления, а также у 7-дневных цыплят-индеек — пульпа селезенки не дифференцирована на красную и белую пульпу. У не? обнаруживаются диффузные лимфоидные скопления и скопления лимфоидных клеток в виде узелков (Рисунок 1. А). Лимфоидная ткань белой пульпы слабо или почти неразвита. Вокруг артерий и артериол выявляется густая сеть, среди них встречаются гемоцитобласты. Паренхима представлена небольшим количеством скоплений незрелых лимфоцитарных элементов, преимущественно эритроцитами, гранулоцитами и в основном псевдоэозинофилами;
- · у цыплят 14-дневного возраста в паренхиме наблюдалось значительное уменьшение количества клеток миелоидного типа, возрастает количество клеток лимфоидного ряда. Паренхима органа незначительно начинает дифференцироваться на красную и белую пульпу. Центральные артерии и многочисленные кисточковые артериолы представляют узкие артериолярные муфты, где наблюдается интенсивный лимфоцитопоэз и более широкие прослойки пульпы, которые в дальнейшем образуют лимфоидные фолликулы и это проявляется скоплением малых лимфоцитов, но их площадь их очень мала. Сформированные лимфоидные фолликулы не встречаются, т. е. лимфоидная ткань В — зависимых зон еще не сформирована (Рисунок Б);
- · у цыплят 21-дневного возраста, в паренхиме органа выявляется значительное уменьшение числа эритроцитов и гранулоцитов. В белой пульпе четко выявляются периартериальная лимфоидная муфта (Т — зависимая зона) и наблюдается увеличение их количества и их площади, она богата клетками, где довольно много лимфоцитов, на разных стадиях развития. В-зависимая зона представлена незначительным количеством диффузной лимфоидной тканью, она наименее развита. В белой пульпе отмечаются несформированные лимфоидные фолликулы без стромы, без четких границ, где они сливаются с красной пульпой;
- · у 30-дневных цыплят-индеек, паренхима органа четко подразделяется на красную и белую пульпу. Количество лимфоидных периваскулярных скоплений расширяется в объеме, отмечается увеличение уровня диффузной инфильтрации лимфоцитами, макрофагами и плазмоцитами. Вокруг кисточковых артериол наблюдается интенсивный лимфоцитопоэз, где довольно много малых лимфоцитов, которое проявляется в Т — зависимой зоне. В белой пульпе, выявляются первые сформированные лимфоидные фолликулы (Взависимая зона). Они чаще располагаются около Т — зависимых зон или артерий мышечного типа. Центры в отдельных лимфоидных фолликулах незначительно разрыхлены, а по периферии клетки лимфоидного ряда располагаются компактно, встречаются отдельные лимфоидные фолликулы в начальной стадии формирования (лимфоидные фолликулы без стромы); (Рисунок.1. В)
Рисунок 1. Селезенка однодневных (А), 14-дневных (Б) и 30-дневных (В) цыплят-индеек.
Окраска гемотоксилином и эозином х 100. А. Недифференцированная лимфоидная ткань пульпы органа. Б. Начало дифференцировки лимфоидной ткани Т — зависимых зон и вокруг артерии и кисточковых артериол. В. Первые сформированные лимфоидные фолликулы белой пульпы органа.
- · у 60-дневных цыплят-индеек, в паренхиме органа гранулоциты отсутствуют, но встречаются сегментоядерные псевдоэозинофилы. Белая пульпа четко отделяется от красной. В белой пульпе разграничена значительное увеличение площади периартериальных лимфоидных муфт и увеличивается число лимфоидных фолликулов, где состав клеток в основном, малыми лимфоцитами, плазмоцитами и макрофагами. Распределение лимфоцитов в лимфоидных фолликулах плотное и равномерное. В функциональном отношении их можно отнести к лимфоидным фолликулам, которые находятся в состоянии относительного покоя. Встречаются лимфоидные фолликулы с герминативным центром; (Рисунок 2. А)
- · у 90-дневного возраста цыплят-индеек, в пульпе органа наблюдается незначительное уменьшение объема количеств Ти Взависимых зон. Такой процесс продолжается до конца 120-дневного возраста. В 120-дневном возрасте Тзависимая зоны имеют более узкие гильзы, Взависимые зоны уменьшаются по числу и размерам. Однако, в 90- и 120-дневном возрасте, в белой пульпе органа, скопление лимфоцитарных элементов плотно, а малые лимфоциты, плазматические клетки и макрофаги встречаются регулярно. Взависимая зона, в сформированном виде наблюдается около периартериальных лимфоидных муфт, и находится в состоянии относительного покоя (Рисунок 2. Б). А в 120-дневном возрасте, встречаются отдельные лимфоидные фолликулы в начальной стадии формирования;
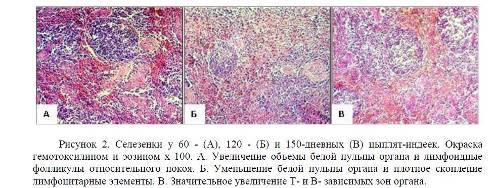
Рисунок 2. Селезенки у 60 — (А), 120 — (Б) и 150-дневных (В) цыплят-индеек.
Окраска гемотоксилином и эозином х 100. А. Увеличение объемы белой пульпы органа и лимфоидные фолликулы относительного покоя. Б. Уменьшение белой пульпы органа и плотное скопление лимфоцитарные элементы. В. Значительное увеличение Ти Взависимых зон органа.
- · у 150-дневных индеек, площадь белой пульпы органа в сравнении с вышесказанными возрастами, значительно увеличивается в объеме и в числе. В белой пульпе четко определяются Ти Взависимые зоны, которые представлены большим количеством зрелыми лимфоцитарными элементами и в основном они состоят из малых лимфоцитов. Лимфоидные фолликулы располагаются ближе к кисточковым артериолам и центральной артерией. В отдельных фолликулах наблюдается плотное скопление лимфоидных клеток без выраженных зон, но выявляются отдельные экземпляры со слабой выраженной герминативными центрами. Это свидетельствует, что в этот период самая высокая степень иммунологической реактивности селезенки, о чем свидетельствуют широкие, хорошо развитые периартериолярные лимфоидные муфты и лимфоидные фолликулы; (Рисунок 2. В)
- · у 180-дневного возраста, скопления клеток в пульпе органа клетки располагаются наиболее рыхло, как в белой, так и в красной. Но, белая пульпа селезенки представлена функционально активными лимфоидными структурами, т. е. Ти В-зависимыми зонами. По сравнений со 150-дневными индейками, где отмечается незначительное уменьшение площади белой пульпы. Ретикулиновая сеть Ти В-зависимых зон разрыхлена, зоны в основном заполнены малыми лимфоцитами. Т — зависимая зона менее широкая, выглядит довольно рыхло, содержат меньше клеток и контуры зон нечеткие. Количество и размеры лимфоидных фолликулов уменьшаются. Отдельные лимфоидные фолликулы со слабо выраженными светлыми центрами, лимфоидные элементы располагаются в центре — рыхло, встречаются фолликулы без герминативных центров (Рисунок 3. А);
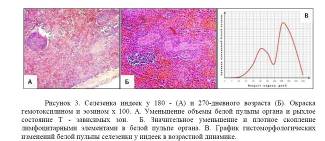
у 270-дневного возраста, наблюдается резкое уменьшение объема периваскулярной лимфоидной ткани. Белая пульпа занимает небольшой объем вокруг кисточковых артериол и пульпарных артерий. Периартериальные лимфоидные муфты узкие, но, сеть ретикулярных волокон Т — зависимой зоны гуще, чем у 180-дневных индеек. Кроме этого, размеры В — зависимых зон белой пульпы уменьшаются почти в двое по сравнению со 180-дневными индейками. Лимфоидные фолликулы представляют собой плотное скопление лимфоцитов, без герминативного центра. Такая картина наблюдаются в белой пульпе 420-дневных индеек. В этом возрасте прослойки белой пульпы сливаются с красной пульпой. В количественном отношении периартериальные лимфоидные муфты значительно уменьшаются. Лимфоидные фолликулы значительно сокращаются по числу и размерами.
У всех возрастов индеек, где есть сформированные лимфоидные фолликулы, они встречаются округлой и округло-овальной или эллипсоидной формы разной величины, со всеми их составляющими и окружены тонкими соединительнотканными волокнами. (Рисунок 3. Б) Обсуждение результатов. На основании проведенных исследований было отмечено, что к моменту вылупления цыплят, процесс дифференцировки, лимфоидной ткани в селезенке еще не закончен. С возрастом паренхима селезенки дифференцируется на белую и красную пульпу. В начале дифференцировки появляется диффузная лимфоидная ткань или Т — зависимая зона, а затем лимфоидные фолликулы или В-зависимая зона. Далее, до 60-дневного возраста, происходит постепенное увеличение объема и количества Ти Взависимых зон, это происходит в основном за счет увеличения числа малых лимфоцитов. После 60-дневного возраста объем Т — зависимой зоны, размеры В-зависимых зон белой пульпы постепенно уменьшаются, но клеточный состав у нее не изменяется. Такой процесс продолжается до 120-дневного возраста. А к 150-дневному возрасту объемы и число Т — и Взависимых зон значительно увеличивается. По сравнению с другими возрастными группами, морфологические параметры белой пульпы у 150-дневных индюшат, достигают максимального значения. Далее, с 420-дневного возраста идет спад, наблюдается постепенное уменьшение белой пульпы, особенно В — зависимой зоны. Тогда, как Т — зависимая зона сохраняется до конца периода. Значительный резкий спад объема белой пульпы наблюдается в 270-дневном возрасте. (Рисунок 3. В) Анализ результатов собственных исследований и литературных источников показал, что к моменту вылупления белая и красная пульпа селезенки цыплят-индеек не полностью дифференцируется. Общий фон микроструктуры белой пульпы селезенки индеек не отличается от микроструктуры белой пульпы селезенки других домашних птиц. Но, возрастная морфофункциональная активность белой пульпы индеек (сроки дифференциация белой пульпы и формирование Т — и В — зависимых зон, начало инволюции белой пульпы и т. д.) отличается от других домашних птиц.
Выводы Таким образом, на основании полученных данных можно сделать вывод о том, что в белой пульпе Т — зависимые зоны формируются к 14-дневному, а Взависимых зонах к 30-дневному возрасту цыплят-индеек. Возрастные морфофункциональные соотношения Ти Взон белой пульпы органа длятся от 30- до 420-дневного возраста, без учета уменьшения их объема и количества. Максимальная морфофункциональная активность Т — и Взависимых зон происходит у — 60- и 150- дневного возраста. Возрастная физиологическая инволюция органа начинается после 150-дневного возраста, а резкий спад наблюдается в 270-дневном возрасте.
- 1. Болотников И. А., Конопатов Ю. В. Физиолого-биохимические основы иммунитета сельскохозяйственной птицы. — Л.: Наука, 1987. — 164 с.
- 2. Селезнев С. Б. Основные принципы структурной организации иммунной системы птиц и млекопитающих. // Морфология и хирургия в практике ветеринарии. — Оренбург. ОГАУ, 1999. — С. 162−164.
- 3. Nagy Z.A. Immunol. //1970. Vj1. 135, p. 653−664.
- 4. Стрельников А. П. Патоморфология и иммуноморфологические реакции у кур при инфекционном бронхите, оспе, колибактериозе и пастерелезе: автореф. дис. д-ра вет. наук: 16.00.02., — М.: МВА, 1987. — 32 с.
- 5. Алдаяров Н. С. Возрастная морфология лимфоидных органов и тканей у кур кыргызской породы: дис. канд. вет. наук: 16.00.02., -Бишкек, 2002. — С. 75−85.
- 6. Асанова Э. И. Морфофункциональная состояние лимфоидных органов и тканей у яков: дис. … канд. биол. наук: 06.02.01., -Бишкек, 2011. — С. 78−85.
- 7. Марасулов А. А. Морфология органов и тканей иммунной системы у кроликов в возрастном аспекте: дис. канд. биол. наук: 06.02.01., -Бишкек, 2011. — С. 66−76.
- 8. Соколов В. И., Чумасов Е. И. Цитология, гистология, эмбриология. — М.:" КолосС", 2004. — С. 247−250.
- 9. Степанова Е. В. Морфология селезенки кур кросса «Хайсекс браун» в постнатальном онтогенезе: дис. канд. вет. наук: 16.00.02., -Брянск, 2006. — 142 с.
- 10. Тубол О. В. Постинкубационный морфогенез селезенки у японских перепелов.// Экологическая безопасность региона: сборник статей Международной научнопрактической конференции.- Брянск, 2009.-С. 21−31.
- 11. Меркулов Г. А. Курс патологогистологической техники. — Л.: Медгиз, 1956. — 262 с.