Индексы видового разнообразия

Шреверо (Schroevers, 1973), встретившись с необходимостью за короткий срок оценить состояние значительного числа водоёмов Дании, применил своеобразный приём, в основу которого положены общие экологические представления. Он принимал, что ненарушенное развитие экосистем ведёт к увеличению их своеобразия, в частности, к увеличению видового разнообразия и стабильности, а последствия антропогенных… Читать ещё >
Индексы видового разнообразия (реферат, курсовая, диплом, контрольная)
В последние время при биологическом анализе вод большое внимание уделяется использованию для этой цели различных индексов применяемых в популяционной экологии. Поскольку с увеличением степени загрязнения видовое разнообразие, как правило, уменьшается, для оценки уровня загрязнения могут быть использованы индексы разнообразия, предложенные разными авторами.
Фишер и др. (Fisher et al., 1943) нашли, что логарифмический ряд хорошо передаёт расположение видов по их численности и предложили константу, а как меру разнообразия:
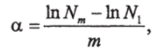
где N— численность вида т в ряду видов, ранжированных по численности;
Nt — численность первого вида с наивысшей численностью; т — порядковый номер вида в ряду 1,2,3…т.
Индекс разнообразия Симпсона (Simpson, 1949):
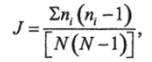
где N—число видов;
п. — число особей /-го вида.
Здесь, чем выше значение индекса, тем меньше видовое разнообразие.
Чрезвычайно широкое применение при изучении природных сообществ получили индексы видового разнообразия, среди которых наиболее популярен индекс Шеннона, заимствованный из теории информации (Shannon, Weaver, 1949):
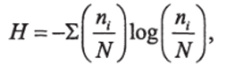
где п, — численность или биомасса /-го вида в сообществе;
N—суммарная численность или биомасса сообщества.
Существует, однако, точка зрения, что индекс Шеннона и другие индексы разнообразия мало пригодны для оценки состояния сообществ (Воробейчик, Садыков, Фарафонтов, 1994). Оказывается, что «здоровой» экосистеме могут соответствовать различные уровни видового разнообразия и в тоже время одно и то же значение индекса может наблюдаться при различных (порой прямо противоположных) состояниях экосистемы.
Маргалеф (Margalef, 1951, 1960), исходя из того, что число видов пропорционально логарифму изученной площади (Gleason, 1922), и считая, что общее число особей пропорционально площади, предложил в качестве меры разнообразия индекс d:
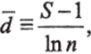
где S — число видов;
In п — натуральный логарифм числа особей.
Индекс d принимает максимальное значение, если все особи принадлежат к разным видам (S = п) и равен нулю, когда все особи принадлежат к одному виду (S = 1). Этот индекс для оценки качества воды по макробентосу использовал Беш (Boesch, 1972), а по деатомеям — Бале (Bahls, 1973).
Позже Маргалеф (1963) предложил другой индекс разнообразия:
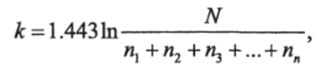
где N— число видов на данном участке;
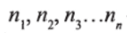 — численносгь отдельных видов на данном участке.
— численносгь отдельных видов на данном участке.
Абсолютная величина индекса возрастает за счёт видов, численность которых наименьшая, то есть, за счёт случайных видов, трофическая значимость которых в данный момент незначительна. Это обстоятельство снижает ценность указанного индекса.
Менхиникк (Menhinick, 1964) рассчитывал видовое разнообразие полевых насекомых по формуле:
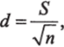
где п — число особей;
5—число видов.
Широкому внедрению биологического анализа в практику изучения степени загрязнения водоёмов препятствует необходимость для его проведения большого количества квалифицированных специалистов по систематике гидробионтов. Для преодоления этого затруднения Кернс и Диксон (1971) предлагают ограничиться установлением количества видов, которые хорошо различимы по форме тела, размеру, цвету и другим признакам. То есть степень загрязнённости определяется по видовому разнообразию и для этого использовать так называемые индексы видового разнообразия. Основанием для этого послужили материалы об уменьшении видового разнообразия организмов при увеличении степени загрязнения водоёмов.
Их исследования показали, что установленное таким образом количество «видов» донных макробеспозвоночных приблизительно равно истинному количеству видов. Для определения индекса последовательного сравнения достаточно, когда в анализируемой пробе находится 200−250 организмов. В незагрязнённых водоёмах индекс имеет значение 12 и выше, а в загрязнённых — 8 и ниже (Макрушин, 1974). Кернс и Диксон предложили индекс последовательного сравнения, который рассчитывается следующим образом:
- 1. Беспозвоночные каждой пробы, отобранные из грунта, перемешиваются в склянке и выливаются в белую эмалированную кювету с плоским дном, на котором нанесены параллельные линии. Комки организмов размываются водой так, чтобы каждая особь лежала отдельно из линий.
- 2. Сравнивают организмы между собой, начиная с первого на первой линии. Сравнивают одновременно лишь два организма: первый со вторым, второй с третьим и т. д. Первый организм обозначается буквой «X». Если второй организм относится к тому же «виду», что и первый, его обозначают буквой «X», если к другому, то буквой «О». Затем сравнивают второй организм с третьим. Если третий организм относится к тому же «виду», что и второй, его обозначают той же буквой, что и второй, если к другому виду, то другой буквой. Для записи рядов используют только две буквы — «X» и «О».
Например, имеется два ряда организмов:
- • АБВАГДЕ — первый ряд;
- • АААБААА — второй ряд.
В этих двух рядах каждая буква обозначает какой-либо вид, название которого неизвестно исследователю, но виды между собой хорошо различимы. Первый ряд записывается буквами.
- • 1 2 3 4 5 6 7;
- • ХОХОХОХ.
Второй ряд записывается буквами:
- • 12 3;
- • XXX О XXX.
- 3. Подсчитывается число пробегов в записанных рядах. Под пробегом понимается ряд одинаковых букв («X» или «О») стоящих рядом. В первом ряду каждая буква — отбельный пробег, во втором — три пробега.
- 4. Подсчитывается промежуточный индекс последовательного сравнения:
П/, = число пробегов в пробе/число особей в пробе.
5. Повторяют определение DJX несколько раз (2−6), предварительно каждый раз перемешав организмы, и подсчитывают среднее значение ?>J: 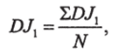
где N — число, показывающее, сколько раз определено DJX.
- 6. Подсчитывают окончательный индекс последовательного сравнения DJ': DJ' = DJ{ X (число «видов» в пробе).
- 7. Определяют 95%-ный интервал достоверности DJ для каждой пробы. Если эти интервалы не перекрываются, значит, имеются различия в видовом разнообразии сравниваемых проб.
Для определения индекса последовательного сравнения достаточно, когда в анализируемой пробе находится 200−250 организмов. Незагрязнённые реки имеют DJ- 12 и выше, в загрязнённых он равен 8 и ниже.
Нередко при биологическом анализе вод большое внимание уделяется использованию для этой цели различных индексов видового разнообразия, применяемых в популяционной биологии. Поскольку с увеличением степени загрязнения видовое разнообразие, как правило, уменьшается, для оценки уровня загрязнения могут быть использованы индексы разнообразия, предложенные разными авторами в разное время.
Согласно большинству предложенных индексов разнообразия, максимум его достигается, когда каждая особь принадлежит к отдельному виду, а минимум — когда все особи относятся к одному виду. Важно, чтобы индексы разнообразия не зависели от размеров пробы. Индексы должны передавать не только распределение видов, но и относительное значение видов в сообществе и, кроме того, быть безразмерными, чему' не удовлетворяют многие предложенные индексы в отличие от индексов, основанных на территории информации.
Арчибальд (Archibald, 1972) при изучении видового разнообразия диатомовых водорослей как показателя качества воды использовал и сравнил между собой пять индексов видового разнообразия:
- • индекс последовательного сравнения Кернса (1968);
- • индекс, основанный на теории информации (Brillouin, 1960);
- • индекс Симпсона (Simpson, 1949);
- • индекс Маргалефа (Margalef, 1951, 1960);
- • индекс Менхиникка (Menhimick, 1964).
Оказалось, что все пять индексов дают одинаковую оценку видового разнообразия. Лучшим из пяти подвергнутых сравнению индексов Арчибальд считает индекс последовательного сравнения Кернса (1968), так как он требует наименьшего количества времени для своего расчёта, а расчёт и истолкование результатов у этого индекса легче, чем у четырёх других.
Симмонс (Simmons, 1972), отмечая преимущества индекса последовательного сравнения (возможность использования его небиологами), указывает на его недостаточную точность, так как некоторые виды и рода может различить только специалист по систематике.
Шреверо (Schroevers, 1973), встретившись с необходимостью за короткий срок оценить состояние значительного числа водоёмов Дании, применил своеобразный приём, в основу которого положены общие экологические представления. Он принимал, что ненарушенное развитие экосистем ведёт к увеличению их своеобразия, в частности, к увеличению видового разнообразия и стабильности, а последствия антропогенных влияний действуют в обратном направлении. Шреверо по результатам обработки проб фитопланктона по четырехбальной системе (1,2, 3, 4) оценивает четыре величины: D — разнообразие, U — своеобразие (unicity), Т — уровень трофии, S — уровень сапробности, и четырьмя разными способами рассчитывает «биологическую оценку». (biolegical value) каждого водоёма, вернее, взятой из него пробы фитопланктона, по формулам:
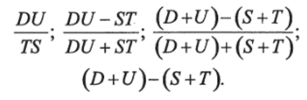
Применив свой приём к пробам из 21 водоёма Дании, автор пришёл к заключению, что, в общем, все четыре способа расчёта приводят к сходному расположению водоёмов в ряду возрастающих значений «биологической оценки».
Общим недостатком всех приведённых выше индексов (за исключением индекса Симпсона) является широкий диапазон получаемых результатов самого индекса, что в существующем виде делает их трудносравнимыми между собой. Последнее устранимо путём приведения их к виду, где шкала индекса изменялась бы от нуля до единицы.
Представляется необходимым, чтобы этому условию отвечали все индексы, имеющие какую-либо экологическую ценность. В такой интерпретации результаты легко представлять в процентах, что сделало бы их удобными в обращении.
Вместе с тем нельзя не отметить, что приведённые выше индексы видового разнообразия, за исключением индекса Шеннона, носят фрагментарный характер. Во всех сообществах имеются доминантные виды, обладающие высокой численностью особей, субдоминантные и случайные виды, число которых достаточно велико, а их численность крайне низкая. Кроме того, характер доминирования претерпевает сезонные пространственные изменения.