Радионуклиды в растениях

Попадая из почвы в растение, радиоактивные элементы в зависимости от свойств проникают в наземные части или же задерживаются в корневой системе. 9°Sr и >37Cs, легко проникают через корневую систему во все части растения, тогда как Се, Ru, Zr, Y, Pu, накапливаются в корневой системе. Распределение радионуклидов в разных частях растений неодинаково: большая часть изотопов концентрируется в листьях… Читать ещё >
Радионуклиды в растениях (реферат, курсовая, диплом, контрольная)
Миграция техногенных радионуклидов в системе «почва-растение» является определяющей в процессах их распределения в наземных экологических системах.
Поступление радионуклидов из почв в растения является первым звеном в пищевой цепочке их перехода из адиабатических компонентов экосистем в биотические, в том числе в организм человека. Попадая из почвы в растение, радиоактивные вещества в зависимости от своих химических свойств проникают в наземные части или же задерживаются в корневой системе. Содержание естественных радионуклидов в растениях зависит от их концентрации в окружающей среде, вида растений, функционального состояния растительности, вида экосистемы и т. д.
Возможны два пути попадания в наземные растения радионуклидов: адсорбция радиоактивных аэрозолей, оседающих из атмосферы, и усвоение (в основном с водой) радионуклидов из почвы. Первый путь представляет собой усвоение радиоактивных аэрозолей надземными органами растений. Так происходит накопление Х*С в процессе фотосинтеза из атмосферного , 4С02. Вторая составляющая связана с корневой системой, которая вместе с восходящими токами воды выносит из почвы растворимые формы радионуклидов. Поэтому степень активности тканей растений определяется не только их видовыми особенностями и концентрацией радионуклидов в питающих горизонтах почвы, но и соотношением в этих горизонтах растворимых обменных фракций с фракциями, находящимися в прочных соединениях с компонентами почвы. На усвоение радионуклидов оказывают влияние возрастные изменения растения, т.к. каждому периоду развития растения отвечает определенный тип физиологических процессов и, следовательно, свой тип питания.
Основными факторами, определяющими уровни накопления радионуклидов в растениях, являются: физико-химические формы радионуклидов, пути поступления и вертикальное распределение радионуклида в почве, агрохимические свойства почв (минеральный состав, кислотность, содержание ионов, вступающих в конкурирующие реакции, ионообменная ёмкость, содержание гуминовых веществ, влажность), особенности метаболизма растений, агротехника выращивания культур.
Коэффициент накопления Кп, Бк/кг/Бк/кг — отношение удельной активности радионуклида в сухом растении Бк/кг к удельной активности 20-см пахотного сухого слоя почвы Бк/кг, на которой выращено растение.
Коэффициент перехода, Кп, Бк/кг/кБк/м2 — отношение удельной активности радионуклида в сухом растении (например, зерне) Бк/кг к плотности загрязнения почвы, на которой выращено растении кБк/м2.
Замечание. Кн у растений всегда выше, чем у животных.
Аэральное поступление важно при радиоактивном загрязнении воздушной среды сразу после радиационного инцидента. Здесь на наземные органы растений оседают радиоактивные аэрозоли, оплавленные силикатные и карбонатные частицы грунта, частицы топлива, высокорадиоактивные «горячие» частицы, входящие в состав «сухого» и «мокрого» выпадения. Растения способны задерживать аэрозоли с размером частиц <45 мкм. При попадании на листья радиоизотопов они проникают во внутреннюю структуру в месте соприкосновения, а затем перемещаются в другие органы растения. Стронций передвигается по растению достаточно быстро и уже через 90 ч. обнаруживается во всех органах растений; ‘-^Cs также обладает очень высокой подвижностью.
Биологическая доступность радионуклидов, образующихся во время ядерного взрыва, определяется его видом. При наземных взрывах на силикатных грунтах частицы локальных выпадений характеризуются слабой растворимостью и низкой биологической доступностью. Радионуклиды локальных выпадений подводных и подземных взрывов, наоборот, обладают высокой биологической доступностью. То же относится к тропои стратосферным выпадениям, когда радиоактивные вещества оседают на поверхность земли в форме мелкодисперсных частиц. При оседании частиц происходит загрязнение надземной части растений выпадающими радионуклидами. Первичное удержание варьируется широко (от нескольких до 95%), что зависит от плотности растительного покрова, морфологических особенностей растений, размеров радиоактивных частиц и метеорологических условий в момент выпадения осадков. Чем больше надземная часть растений, тем больше величина первичного удержания. Так, например, в период максимального развития надземной части пшеница удерживает до 70% водорастворимых радионуклидов, горох — до 75%, ячмень, овес и просо — до 50%, картофель — до 25%, трава -25%. По степени задерживания радионуклидов растения располагаются в ряд: капуста>свёкла> картофель> пшеница>травяная растительность (за исключением мхов, лишайников, грибов, которые отличаются высокой степенью задержки радионуклидов). Нерастворимые радионуклиды загрязняют растения только с поверхности, а растворимые поглощаются через листья, стебли и плоды. По скорости листовой абсорбции радионуклиды располагаются в ряду: Cs>Ba>>Ru. Для травянистой растительности у 9°Sr и wCs коэффициент первичного удержания меняется в диапазоне 204−40 (Бк/кг)/(Бк/м2-сут.).
Первичное удержание и последующие процессы потерь радиоактивности зависят от размера частиц и вида выпадений, площади удерживающей поверхности и плотности растительного покрова, морфологии растений и типа травостоя, урожайности наземной массы, метеоусловий во время и после выпадения радиоактивных осадков и др. По мере увеличения запаса растительной массы на единицу площади повышается степень удерживания радионуклидов. Первичное удержание водорастворимых форм радиоактивных веществ в 44−7 раз выше, чем нерастворимых. С увеличением размера частиц уменьшается их удерживание растениями. Осевшие на надземных частях растений радионуклиды инкорпорируются во внутренние ткани в результате биологических процессов и накапливаются в растениях. Процесс накопления радионуклидов зависит от стадии развития растения, а она в свою очередь от времени года.
Осевшие на поверхности растения радионуклиды могут смываться дождем, сдуваться ветром или поступать во внутренние части растения и усваиваться. Мелкие частицы и водорастворимые формы закрепляются в 44−7 раз прочнее, чем крупные и твёрдые нерастворимые частицы. Максимальные потери радиоактивности происходят в первые 24−3 суток; за 7 суток она снижается на 8о%. Потери фиксированных радионуклидов мало зависят от погодных условий и определяются свойствами радионуклидов и биологическими особенностями растений.
Значимость аэрального пути поступления радионуклидов не зависит от периода полураспада радионуклида, а корневое поступление существенно лишь для относительно долгоживущих радионуклидов, не успевающих распасться до усвоения их корнями растений. Внекорневое накопление радионуклидов может происходить двумя путями: при выпадении радионуклидов из атмосферы непосредственно на растительность и отложение на растительности радионуклидов, попавших первоначально на почву, а затем за счёт вторичного пылеобразования осевших на растительности.
Для долгоживущих радионуклидов с течением времени после выпадения радионуклидов на почву и их миграции в почве к корневой системе растений возрастает роль накопления радионуклидов через корни растений. Коэффициент накопления сильно меняется в зависимости от почвы, вида растительности и условий окружающей среды. С течением времени он может увеличиваться по мере достижения радионуклидом оптимального положения относительно корневой системы, затем уменьшаться при изменении физико-химического состояния радионуклида и перехода радионуклида в почву ниже корневой системы.
Техногенные радионуклиды обычно находятся в соединениях доступных для усвоения растениями. Однако с течением времени они с различной скоростью переходят в трудно растворимые формы. Так, 137Cs в почвах довольно быстро переходит в труднодоступную форму, тогда как 9°Sr может длительно (многие годы) оставаться в обменной форме. ^°Sr хорошо усваивается растениями, причём его накопление в них обратно пропорционально количеству обменного кальция в почве. В подвижной форме 137Cs также хорошо усваивается растениями.
Коэффициенты накопления снижаются с течением времени, что связано с миграцией радионуклидов за пределы корнеобитаемого слоя и увеличением энергии связи соединений с почвенным поглощающим комплексом. В многолетней динамике корневого поступления радионуклидов выделяют два периода. Период интенсивного корневого накопления имеет место в первые вегетационные сезоны после выпадений, что обусловлено нахождением в поверхностном слое почвы. Здесь биологическая доступность радионуклидов максимальна. Затем следует период установления квазиравновесного распределения радионуклида в системе почва-растение управляемый установлением постоянного распределения радионуклидов между твёрдой и жидкой фазой почвы.
При выпадении радиоактивных веществ на луговую и пастбищную растительность значительная их часть задерживается в нижней части растений и в верхнем слое прикорневой дернины, откуда поступают в растения через основание стебля и поверхностные корни, также образуют «дернинный резервуар». Радиоактивные вещества, попадающие в почву, частично вымываются и загрязняют грунтовые воды, но почва прочно удерживает их и обеспечивает их нахождение в почвенном горизонте и поступление в растения за счёт почвенного поглощающего комплекса.
Содержание радионуклидов в растениях зависит от их индивидуальной способности избирательно аккумулировать в себе определенные химические элементы. К таким растениям относятся мхи и лишайники. Особенности лишайников (многолетний характер роста, воздушный тип питания растений) обусловливает их высокие уровни радиоактивного загрязнения. Ягель — биоиндикатор, отражающий состояние окружающей среды. Поэтому мохово-лишайниковую растительность используют при проведении радиоэкологического мониторинга окружающей среды.
Замечание 1. Биологическое накопление свойственно растениям, которые, аккумулируя определенные химические элементы, изменяют окраску хвои, листьев, цветков и плодов. Это служит, индикаторным, признаком, при поисках полезных ископаемых. Например, береза и осина в Восточной Сибири накапливает в своей древесине значительные, содержания 9Sr, что приводит к появлению необычной окраски — неестественно зелёного цвета. В ареале рассеяния урановых месторождений лепестки иван-чая вместо розовых становятся белыми и ярко-пурпуровыми, у голубики плоды вместо темно-синих становятся белыми и т. д.
Замечание 2. Основная часть , 37Сs во мхах и лишайниках обязана своим присутствием радиоактивным осадкам непосредственно из атмосферы. Поэтому использовать эти растения как индикаторы перехода радиоактивности из грунта некорректно. Грибы создают обширную сеть мицелия, который эффективно экстрагирует из почвы микроэлементы, концентрируя их в плодовых телах. При этом мицеллы грибов разрушают горячие частицы, вовлекая радионуклиды в биоту. Можно использовать грибы в качестве биоиндикаторов радиоактивного загрязнения и определить плотность загрязнения почвы Cs по данным о загрязнении грибов.
Некоторые виды лекарственных растений (крапива двудомная, череда трёхраздельная) накапливают значительные количества 9°Sr и wCs. Причём, если крапива накапливает оба радионуклида, то череда — преимущественно *37Cs. Сильно поглощают радионуклиды сосна, береза, ель, осина, рябина, малина, черника, укроп, клюква, петрушка, шпинат, бобовые, злаки, гречка, рапс, ромашка, мхи, смородина и др. Много Th откладывается в листьях осины, черемухи и пихты. Меньше поглощают радионуклиды ольха, фруктовые деревья, капуста, огурцы, картофель, кабачки, томаты, лук, перец, чеснок, свёкла, морковь, редис, хрен, ирис, редька.
По уровню накопления радионуклидов огородные культуры располагаются в порядке: огурцы>томаты> капуста (кольраби>цветная>ранняя жраснокочанная). При возделывании картофеля наименьшее загрязнение клубней наблюдается у сортов Аксамит, Сантэ, Синтез. Из плодово-ягодных культур больше накапливают радионуклиды ягоды красной и чёрной смородины, крыжовник, меньше — земляника садовая, белая смородина, клубника, малина, яблоки, груши, вишни, сливы, черешня. Наименьшее загрязнение имеют ягоды рябины, земляники, малины, а наибольшее — черника, клюква, голубика, брусника.
OIC4CI OlCNCI.
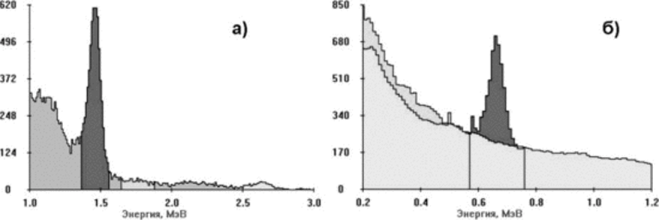
Рис. 15. у-Спектры природных объектов: а — гранитная крошка с Карельского перешейка (хорошо видна линия «°К, присутствуют линии Ra и Th); б — сухие грибы из-под Луги (видна линия wcs).
Сельскохозяйственные растения меньше загрязнены радионуклидами, чем дикорастутцие, так как производится обработка почвы, вносятся удобрения и т. п. У некоторых сельскохозяйственных растений хозяйственно ценные части урожая достаточно надежно защищены от загрязнения — зернобобовых культур, просо, риса, ячменя, овса, початки кукурузы, клубни картофеля, корнеплоды.
Чем древнее по своему происхождению растения, тем большей способностью к накоплению радиоактивного вещества они обладают. Например, у папоротников радиоактивность ниже, чем у лишайников. Ещё более низкую радиоактивность имеют голосеменные растения, и совсем незначительную — цветковые. В ходе эволюции растения выработали приспособления для уменьшения накопления в своих тканях радионуклидов, которые могут вызывать неблагоприятные изменения. Культурные растения — сахарная свекла, пшеница — по сравнению со своими дикими предками содержат меньше радиоактивных элементов.
Попадая из почвы в растение, радиоактивные элементы в зависимости от свойств проникают в наземные части или же задерживаются в корневой системе. 9°Sr и >37Cs, легко проникают через корневую систему во все части растения, тогда как Се, Ru, Zr, Y, Pu, накапливаются в корневой системе. Распределение радионуклидов в разных частях растений неодинаково: большая часть изотопов концентрируется в листьях, стеблях и корнях, меньше — в соцветиях и еще меньше — в плодах и семенах. Исключением является изотоп wCs, концентрация которого в семенах может достигать ю% от его содержания в надземной части растений. В древесине деревьев накапливается меньше радионуклидов, чем в листьях или хвое (i/6 находится в стволе, 5/6 — в коре, ветвях и листьях). Во фруктах они находятся в косточках, в капусте — в верхних листах и кочерыжке, в свёкле и моркови — в начале ботвы и т. п. Кп у молодых деревьев выше, чем у старых, поскольку у последних обменные процессы замедленны. Больше радионуклидов накапливают многолетние луговые травы, чем однолетние сельскохозяйственные культуры. Накопление радионуклидов зависит от площади поверхности растения и его строения. Так, форма соцветий пшеницы способствует удержанию выпадающих мельчайших аэрозолей. В лесу больше всего задерживают радионуклиды хвойные породы деревьев, что связано с медленной сменой хвои. Лиственные породы деревьев ежегодно сбрасывают свой покров, поэтому степень накопления у них меньше.
Повышенное содержание U наблюдается в корнях и отличается от содержания нуклидов в надземной части в 10,5 раз. В листьях и стеблях исследуемых растений содержится до 12 Б к/кг U, в то время как в корнях — до 65 Бк/кг (клевер красный). Для Ат разница между содержанием нуклида в подземной и надземной части растения достигает -1,5 раз. Более высокое значение содержания Ат в корнях отмечено для клевера розового и люцерны, минимальное — горчицы белой. С повышением кислотности почвы, как для U, так и для Ат в почвенном растворе увеличивается общее количество водорастворимых и обменных форм на 32% (для изотопов U) и на 34,7% Для Ат. В то же время содержание водорастворимых форм, доступных для растений, возрастает на 63,2% для изотопов U и в 2,71 раза для Ат. Изотопы U и Ат характеризуются малой подвижностью в системе «почва-растение». Кп в растениях Ат на 1н-2 порядка ниже по сравнению с Кн 137Cs или 9°Sr. Накопление растениями изотопов U и Ат уменьшается с уменьшением значения pH, гумуса и обменного Са.
Для радионуклидов, поступающих по корневому пути свойственно неравномерное распределение в растениях: радиоактивные Sr накапливается в надземных частях растений; Cs — распределяется равномерно; РЗЭ и ТУЭ — в корневых системах. В ходе вегетации абсолютное количество радиоизотопов в растениях возрастает, а относительное содержание на единицу массы сухого вещества снижается. С увеличением урожайности уменьшается относительная концентрация радионуклидов.
Накопление радиоактивных элементов растениями зависит от содержания их в почве, от свойств почв (pH, гранулометрический состав, гумус и др.), их доступности, вида растения, фазы его развития, природноклиматических условий и др. Концентрация радионуклидов в растениях зависит от таких факторов, как концентрации и формы их нахождения в корнеобитаемом слое почвы; геохимические особенности радионуклидов и присутствие близких по химическим свойствам элементов (носителей); видовые особенности растения. Гранулометрический состав почвы влияет на закрепление радиоактивных веществ в почве: тяжёлые почвы сильнее закрепляют радионуклиды, чем лёгкие. Важен и минералогический состав почвы. Наибольшей поглотительной способностью обладают минералы монтмориллонитовой группы и гидрослюды, наименьшей — каолиниты.
Много радионуклидов содержится в торфяниках, меньше — в песках и ещё меньше — в супеси и суглинке. Повышенное количество радионуклидов находится в растениях переувлажнённых и лугопастбищных угодий. Сорбция на гумусовом веществе с последующим переходом в необменные формы делает радионуклиды слабо доступными для растений, тем более, что гумусные кислоты растворяются лишь в щелочах. На тех почвах, где радионуклиды прочно связана в гумусных горизонтах, наблюдается снижение Кн• Коэффициенты перехода 9 $г и ‘З/Cs в сельскохозяйственные культуры снижаются при увеличении ёмкости катионного обмена почв, содержания гумуса, элементов минерального питания, а также при снижении кислотности почвенного раствора. Максимальные коэффициенты перехода наблюдаются для торфяных почв и минеральных почв лёгкого гранулометрического состава, а минимальные — для плодородных тяжёлосуглинистых и глинистых почв (дерново-подзолистые, серые лесные, чернозёмы). На поведение радионуклида в системе почва-растения оказывает влияние концентрация его носителя: для wCs — стабильный Cs и К, для 9°Sr — стабильный Sr и Са. Увеличение концентрации носителя приводит к снижению перехода радионуклида в растения. Так, при увеличения уровня увлажнения величина переходаCs в растительность возрастает в боо раз. Чем прочнее радиоизотоп фиксируется в почве, тем меньшее его количество попадает в растение. Например, овёс, выращенный на песке, накапливает 9°Sr в несколько раз больше, чем овёс, произрастающий на тяжёлом суглинке. При этом из глинистого песка поступает ~ю%, а из тяжёлого суглинка — 1% всего 9°Sr, внесённого в почву. Накопление растениями элементов из почв образует ряд: Sr>J>Ba>>Ce>Y, Pm, Zr, Nb>Pu.
На корневое поступление радионуклидов влияют агрохимические характеристики почвы: с увеличением доли глины и ила в твёрдой фазе почвы повышается доля вторичных минералов, определяющих значительные ёмкость почвенного поглощающего комплекса, увеличивается необменная сорбция радионуклидов и снижается их корневое поступление; увеличение влажности почвы повышает переход радионуклидов в растения; повышение кислотности почвенного раствора увеличивает биологическую доступность Cs и Sr; повышение доли органического вещества в почвах снижает корневое поступление радионуклидов в растения. Наименьший переход радионуклидов в растения наблюдается в регионах, где преобладают чернозёмные почвы, наибольший — в регионах с торфяноболотистыми почвами. Высокие коэффициенты перехода радионуклидов характерны также для песчаных почв. При повышении содержания гумуса в почве от 1 до 3,5% переход радионуклидов в растения снижается в 2 раза.
Минимальное накопление радионуклидов в растениеводческой продукции наблюдается при оптимальных показателях кислотности почв (pH), которые для дерново-подзолистых почв составляют: глинистых и суглинистых — 6,04−6,7; супесчаных — 5,84−6,2; песчаных — 5,64−5,8. На торфяноболотных и минеральных почвах сенокосов и пастбищ оптимальные показатели pH составляют соответственно 5,04−5,3 и 5,84−6,2. На песках переход радионуклидов в растения вдвое выше, чем на суглинках, особенно при низкой обеспеченности почв обменным калием. На переувлажненных песчаных почвах высокая степень загрязнения травяных кормов наблюдается даже при относительно низких плотностях загрязнения почв радионуклидами, а на окультуренных участках лессовидных и моренных суглинков возможно получение продукции с допустимым содержанием ‘37Cs при плотности загрязнения до 7404−1110 кБк/м2.
Существует обратная корреляция между количеством обменного К в почвах и накоплением *47Cs в растениях. Калийные удобрения снижают поступление wCs в растения, причём этот процесс происходит интенсивнее при оптимизации содержания в почве наиболее значимых микроэлементов (Мо, В, Си и др). Органические удобрения, особенно торф с низким pH, вызывают повышение доступности для растений *37Cs, этот негативный эффект торфа компенсирует известкование почвы.
Коэффициенты накопления и перехода 9°Sr из почвы в зерно ржи, овса и озимой пшеницы лежат в диапазоне 0,044−2,0 Бк/кг/Бк/кг и 0,154−5,5 Бк/кг/кБк/м2. Медианные значения коэффициентов перехода (А*п, Бк/кг/кБк/м2) 9°Sr из почвы в зерно более чем в ю раз превышают Кц >37Cs. Для зерна озимой ржи, пшеницы и овса имеет место обратно пропорциональная зависимость Кп 9°Sr от содержания в почве обменного кальция. Известкование кислых дерново-подзолистых, песчаных и супесчаных почв, внесение фосфорных удобрений в 3 раза уменьшает содержание 90Sr. Доля 9°Sr в обменной форме возрастает с увеличением кислотности почвы.
Динамика загрязнения 9°Sr растительности определяется кинетикой растворения топливных частиц (ТЧ), вертикальной миграцией 9°Sr и обусловленным этим изменением содержания мобильного 9°Sr в корнеобитаемом слое. В зависимости от скорости растворения ТЧ корневое загрязнение 9°Sr растительности в первые годы после аварии растёт и достигает максимума на 24−30 год в зависимости от скорости растворения ТЧ, обусловленной степенью окисления матрицы частиц и кислотностью среды. Затем содержание в зернеSr уменьшается из-за его радиоактивного распада и вертикальной миграции из пахотного корнеобитаемого слоя.
В настоящее время основная масса топливных частиц растворилась и 404−90% 9°Sr в дерново-подзолистых почвах легкого механического состава уже находится в обменной форме. Удельная активность 9°Sr в растениях за послеаварийный период достигла своего максимума, и на протяжении последних лет наблюдается монотонное уменьшение содержанияSr в зерне. Загрязненное зерно используют при производстве спирта, в качестве семенного материала и на корм скоту.
Радионуклиды переводятся из почвы в растения, дающие пышную зеленую массу. Повышение температуры воды, используемой для полива, несколько увеличивает миграцию радионуклидов из почвы в растения. В сезонной динамике хз?Cs в ассимилирующих органах древесных пород отмечается однонаправленное снижение его концентрации от весны к осени, a 9°Sr, напротив, — рост рассматриваемого показателя в период вегетации. В то же время в древесине минимум концентрации wCs приурочен к началу интенсивного весеннего сокодвижения. Внесение в почву Fe, Со и Y совместно с комплексоном ЭДТА повышает подвижность Sr и Cs и увеличивает поступления их в растения.
Переход 238U из почвы в растение определяется особенностями его поведения в системах твёрдая фаза почвы — почвенный раствор и почвенный раствор — растение. Для усвоения корневыми системами растений доступна только та часть ионов радионуклида или элементов минерального питания, которая способна перейти в раствор из сорбированного твёрдой фазой почвы состояния. Доступность 238U растениям определяется следующим рядом исходных химических форм радионуклида: (NH4)2U207>
uo2ci2>uo3>u3o8> U02S04>U02.
Наиболее высокие значения коэффициентов биологического поглощения растениями U отмечаются для районов горной тундры и средней тайги (о, 21 и 0,17 соответственно), где в почвах преобладают минеральнообломочные формы U. Для горно-тундровых и горнолесных ландшафтов района северной тайги среднее значение Кн существенно ниже (о, о8). Для этого района в большей степени была отмечена аккумуляция U гумусовым веществом и интенсивная водная его миграция. Самые низкие значения Ки характерны для района лесостепи (0,005), где процессы аккумуляции U почвами имеют особенно большое значение.
Коэффициент накопления U сельскохозяйственными растениями зависит от типа почвы, формы удобрений, вида растений и составляет 0,0001*0,1. Низшие растения (мхи и лишайники) накапливают большие концентрации U по сравнению с высшими растениями. Довольно высоки коэффициенты перехода U у горчицы белой и рапса ярового. Среди высших растений не обнаружено концентраторов U (исключение — астрагал). При кларковом содержании U в почвах (п-ю6 г/г) концентрация его в золе растений составляет 5−10−7 г/г; при увеличении концентрации U в почвах до п-юУп-ю-з г/г его содержание в золе растений увеличивается, но не пропорционально содержанию в почве. Это связано с существованием у растений «физиологического барьера», препятствующего поглощению больших количеств U. Роль такого барьера играют корни и отчасти листья.
Переход 2з"и в растительность варьирует в пределах 2*13 раз и определяется строением корневой системы. Корни растений выделяют ионы водорода, аминокислоты и хелаты, влияющие на усвоение многих элементов. Глубина размещения корней различных видов растений играет важную роль в извлечении радиоактивных элементов из почвы. Число придаточных корней увеличивается в ряду: корневищные бобовые<�разнотравье <�злаковые, ряд характеризующий переход 238U в луговые растения следующий: злаковые> разнотравье>бобовые. В золе растений (при содержании U в почве Ю'4%) его концентрация составляет 1,5−10−5%. Концентрации U в высушенных растениях варьируются в пределах 5*60 ррт> а зола от сожжённых растений содержит до 4 ppm U. Содержание U в золе растений зависит от их видовой принадлежности, возраста и органа. Аккумуляция U в наземной части растений идёт в вегетационных органах. В вегетативной массе зернобобовых (соя, нут) и зерновых (пшеница, ячмень) культур содержится 238U в 2,44−4,2 раза больше, чем в генеративных органах. Максимальная концентрация 2з8и отмечена в листьях чая и вегетативной массе зернобобовых культур, а минимальная — в генеративной части зерновых. По степени аккумуляции 2з8и можно составить ряд: чай>соя>нут> хлопчатник>пшеница>ячмень. Большое накопление U отмечается у многолетних древесных и кустарниковых растений. Прекрасными аккумуляторами U являются мхи (з*Ю'3 г/кг золы), грибы и водоросли. Грибы переводят U в довольно безвредные химические соединения.
На снижение накопления U растениями влияет применение органических удобрений (особенно навоза) и извести. Снижение концентрации радионуклидов в урожае при внесении удобрений обусловлено такими причинами, как увеличение биомассы («разбавление» радионуклидов); повышение концентрации в почве обменных катионов, усиление антагонизма между ионами радионуклидов и ионами вносимых солей при корневом усвоении; изменение доступности для корневых систем радионуклидов вследствие перевода их в труднодоступные соединения и обменной фиксации в результате реакции радионуклидов с вносимым удобрением.
Чем выше Кн U, тем меньше коэффициент водной миграции Къод.
В районах, где U изоморфно входит в минералы РЗЭ и слабо подвижен, /Свод имеют минимальные значения. В то же время U здесь доступен для растений, так как величины его Кн максимальны. В таёжном районе преобладают рассеянные формы U в подстилающих гранитах. Уран сравнительно легко извлекается природными водами из обломков пород, а затем прочно связывается гумусом на восстановительных барьерах и становится слабо доступным для растений. Здесь Кц U низкие.
В природных условиях в окислительной обстановке U является сильным мигрантом, но считается металлом слабого биологического поглощения. Доступными для растений формами U являются U-протеиновые комплексы. Кн U растениями <0,40. В растениях U концентрируется производными целлюлозы.
Среднее содержание U для наземной растительности 38 мкг/кг (0,46 Бк/кг) в интервале от 5 до 69 мкг/кг (0,034−0,84 Бк/кг). Для древесной растительности Сибири активность 2з8и составляет (Бк/кг золы): берёза бородавчатая — 12,21, сосна обыкновенная — 12,21, осина дрожащая — 3,66, лиственницы даурская и сибирская — 122,10. В злаковых культурах U накапливается в вегетативной массе. Коэффициенты накопления U в зерне и соломе озимой пшеницы на разных типах почв колеблются в интервалах 0,44−0,003 и 0,0024−0,006 соответственно. Коэффициенты накопления урана кукурузой 0,14−0,012. Радиоактивность травы равна мо^-ю, 5−10−9 Ки/кг, мхов — o, 5*iO'74-mo-7. Максимальные концентрации U отмечены в ромашке аптечной (35,41 Бк/кг). Содержание U в разных видах полыни варьирует от 5,01 до 18,32 Бк/кг. Сильно накапливают U гречишные (15,47 Бк/кг). Не обнаружен (аз8и в надземной массе растений «10,46, в корнях — 14,55 Бк/кг.
При избыточном содержании U в почве у растений наблюдается карликовость, а также изменяется окраска и форма органов: розовые цветки Иван-чая изменяют окраску на белую или ярко-пурпурную, а у голубики вместо тёмно-синих формируются белые или зеленоватые плоды неправильной формы. Изотопное отношение 234U/238U зависит от формы нахождения дочернего изотопа и может служить её трассером. При гомогенном распределении всех изотопов U в воде их изотопное отношение в растении будет таким же, как и в питающей среде. Если более подвижный 234U из почвенных растворов прочно фиксирован гумусом, он является менее доступным для растений, чем 2з8и, преимущественно находящийся в минеральной матрице почв. В этом случае (2:ии/2з8и)раСТ<(234и/2з8и)почв.
Уран не является биогенным элементом: Кц< 1. При этом наблюдается зависимость Кн от содержания U в почвах. При содержаниях U, близких к ю-3%, Кн уменьшается. Это объясняется существованием в корнях растений биологического барьера, препятствующего проникновению в надземную часть концентраций выше нескольких миллиграммов на 1 кг золы. Поэтому в корнях накапливается больше U, чем в надземных органах. Уран помимо корней накапливается в более старых органах (древесине, крупных ветках, коре), а не в молодых листьях и хвое. Для Полярного Урала Кн U для мхов достигает 6,9 при среднем значении 2,3, в то время как для кустарников эта величина не превышает 1. Низшие растения концентрируют U в большей степени, чем высшие. Из древесной растительности наиболее высокие Кн отмечены у березовых (Кн для березы пушистой «о, 9 при среднем значении для всех видов растительности *0,05).
Минимальное накопление U отмечается у травянистой растительности, особенно у злаковых. У культурных зерновых растений зерно содержит в 2 раза меньше U, чем солома. Удельная активность изотопов U в продуктах питания составляет десятые доли-единицы Бк/кг. Содержание 226Ra в растениях и пищевых продуктах варьируется в интервале 3,7−5-23 Бк/кг. Содержание Ra в зерне 2,3-ю15 Ки/г, в картофеле 9,6-ю15, в сухом молоке 1,7-ю15, в треске 4,0-ю15, моркови и капусте 1,7-ю15 Ки/г. Содержание 226Ra в теле человека колеблется в пределах (34−1400) пКи/г, 228Ra 50 пКи, а содержание 2ЮРЬ и находящегося с ним в равновесии 21°Bi — 744−185 Бк/кг. 210Ро содержится в растительных и животных организмах в количествах, равных 504−100% от содержания 2ЮРЬ. Важно поступление 210РЬ и 2ЮРо в табачные растения из почвы. В лишайниках тундры содержится 2К,Ро до 2000 Бк/кг, в чайном листе — >15 Бк/кг, в табаках — 55 Бк/кг.
Растения плохо накапливают U — сказываются барьеры, ограничивающие его поступление в клетки. По отношению к Ra таких барьеров нет: он концентрируется в молодых частях растений, в том числе в листьях и цветках, но у некоторых видов — в корнях. Радий обнаружен в травянистых и цветочных растениях, кофе, древесине, морских водорослях, мхах и т. д. Бразильские орехи содержат 404−260 Бк/кг Ra, что в юоо раз выше, чем в других продуктах питания. Кн Ra для районов тундры и тайги выше, чем для чернозёмных лесостепных районов. В первом случае Ra связан в почвах по механизму ионного обмена, а в последнем — прочно фиксирован в слаборастворимых гуматах и сульфатах Са и Ва.
В корнях и листьях травянистых растений Ra больше, чем в стеблях и органах размножения; больше всего Ra в коре и древесине. Среднее содержание Ra в цветковых растениях (0,34−9,0)-ю*11 Ки/кг, в морских водорослях (0,24−3,2)-ю11 Ки/кг. В корнях и листьях травянистых растений Ra больше, чем в стеблях и органах размножения; больше всего Ra в коре и древесине. Содержание Ra в цветковых растениях (о, з+9,о)-юп Ки/кг, в морских водорослях (о, 2+з, 2>10‘11 Ки/кг, в пшенице концентрация Ra (го+гбИо*^ г/г, в картофеле (67+125)-ю^ г/г, в мясе 8 io ,5 г/г.
Радон не вступает в химические связи с другими элементами, но сравнительно хорошо растворим в воде и способен мигрировать на значительные расстояния, что создаёт благоприятные условия для рассеяния в биосфере долгоживущих продуктов его распада — 210РЬ (22 л) и 210Ро (140 дн). Главным путём загрязнения растительности 21°РЬ является непосредственное отложение его на поверхности наземных частей растения с аэрозолями (-85%), второй путь загрязнения — переход в растение через корневую систему из почвы менее значим (-15%). Его содержание в траве определяется количеством осадков, а не содержанием в почве. 2ЮРЬ наиболее сильно аккумулируется лишайниками (содержание в 50 раз выше, чем в высших растениях) и мхами (в 30 раз выше). 2ЮРЬ хорошо усваивается растительными организмами. Есть корреляция содержания 2ЮРЬ в поверхностном слое почвы с биомассой растения. Лишайники не имеют корневой системы, поэтому РЬ и Ро поступают в них из воздуха. Большая сорбционная способность фитомассы лишайников к содержащимся в воздухе РЬ и Ро и большая продолжительность жизни лишайников (>300 л) определяют в них высокую концентрацию этих элементов. Период полуочищения лишайников от РЬ и Ро «7 л.
Табл. 20. Содержание 4°К в основных продуктах питания человека.
Продукт. | Содержание калия. | Удельная активность, Бк/кг. |
Пшеница. | 0,5. | |
Рожь. | 0,6. | |
Горох. | 0,9. | |
Крупа гречневая. | 0,13. | |
Картофель. | о, 45. | |
Морковь. | 0,29. | *4. |
Лук. | 0,15. | 44,4. |
Огурцы. | 0,34. | |
Лимоны. | 07. | |
Клюква. | 0,15. | 44,4. |
Говядина. | 0,37. | |
Свинина. | 0,1. | 33,3. |
Рыба. | 0,26. | 77,7. |
Икра. | 0,42. | |
Молоко коровье. | 0,14. | 44,4. |
Масло сливочное. | 0,014. |
Наилучшими аккумуляторами 2^2ТЬ, содержащегося в поверхностном слое почвы, являются черника, вереск и лишайники. Среди пищевых растений наиболее активно накапливают ТЬ помидоры, далее следуют огурцы, сахарная свекла, горох, капуста и столовая свёкла.
В растениях калий локализуется в паренхиме, в точках роста, в сетчатых трубках, а также во вторичных тканях — древесине, коре. Он принимает участие в углеводном и белковом обмене растений. Содержание естественной смеси калия составляет в пищевых продуктах 0,1+3%, что соответствует значениям удельной активности веществ по *°К от 30+900 Бк/кг. Особо богаты 4°К: фасоль — 299, грибы — 227, картофель — 174, орехи — 210, клюква — 355 Бк/кг. Содержание 4°К в большинстве продуктов питания составляет от одного до нескольких единиц-десятков Бк/кг, повышаясь до 340740 Бк/кг у бобовых и снижаясь до единиц Бк/кг у рафинированных продуктов (шлифованный рис, пшеничная мука высшего сорта и изделия из нее и другие). 4"к особенно интенсивно усваивается фасолью (229 Бк/кг), картофелем (174 Бк/кг), орехами (210 Бк/кг), клюквой (355 Бк/кг). Минимальные концентрации 4к отмечаются в рисе, бруснике — 8-н8 Бк/кг, а максимальные в клюкве, грибах — 280350 Бк/кг.
Из почвы растения поглощают лишь те радиоактивные вещества, которые растворяются в воде. По степени поступления из почвы они образуют ряд: 8^9oSr>131J>14°Ba>137Cs>lo6Ru>144Ce>9°Y>45Zn>95Nb>210Po. По интегральной аккумуляции 9osr> wCs и 210Pb образуют ряд: сено естественных трав>сено сеяных трав>солома>трава естественных пастбищ=силос>сенаж > комбикорм>трава сеяная>зернофураж>морковь>свёклажартофель.
В растения 9"Sr послушает непосредственно через листья или из почвы через корни. Поглощение в листьях идёт медленно, а отложенный изотоп легко сдувается с них ветром и смывается дождями. Количество 9°Sr, переходящее из листьев или с поверхности зёрен в другие части, растения мала. Поглощение из почвы — основной путь поступления 9Sr в растения. Поглощение велико из почв с низким содержанием Са, при добавлении извести оно снижается. На корневое поглощение 9Sr влияет наличие глины и количество гумуса в почве, pH, концентрации электролитов и влагосодержание. Добавление органических веществ и удобрений в почву влияет на поглощения 9°Sr растениями, но довольно слабо.
Для многолетней динамики 9"Sr характерно нарастание корневого потребления до определенного уровня, затем некоторая стабилизация его содержания в растениях и последующее снижение за счёт радиоактивного распада и необменного закрепления в почве. 9"Sr интенсивнее поглощается растениями из лёгких песчаных почв с низким значением pH, бедных органическими веществами и кальцием. Легче всего процесс перехода 9Sr в растения идёт из дерново-подзолистых почв, далее следуют серозёмы и каштановые тёмные карбонатные.
Злаковыми растениями поглощение 9ogr идёт эффективно, что приводит к высоким концентрациям 9"Sr в зерне и траве пастбищ в ранние периоды после осаждения 9Sr. Из почвы через корневую систему 9Sr поступает в растения и входит в состав зерна, бобов, моркови и друтих продуктов (на почвах с невысоким уровнем загрязнения 9°Sr (0,1 Ки/км2) наименьшее его количество обнаружено в горохе, а наибольшее — в сое).
Наиболее высоким содержанием 9"Sr на единицу массы отличаются вегетативные органы растений (десятки и сотни раз выше, чем в зерне, клубнях и корнеплодах); в зерне гречихи наблюдается максимальная концентрация, минимальная — в зерне гороха. Относительно большие количества 9"Sr накапливают бобовые, клубни и корнеплоды, злаки. Коэффициенты накопления и перехода 9°Sr из почвы в зерно обратнопропорционально зависят от содержания в почве обменного кальция. Хотя 9"Sr слабо передвигается внутрь растения при попадании его на листья, для овощных культур (капуста, томаты, огурцы, пищевая зелень и т. д.) это опасно.
В миграции 9osr большую роль играет лесная растительность. Задержанные поверхностью листьев и хвои радионуклиды поступают на поверхность почвы с опавшими листьями и хвоей. В лиственных подстилках содержание 9°Sr постепенно падает от верхнего слоя к нижнему, в хвойных происходит значительное накопление радионуклида в нижней гумусированной части подстилки. 9"Sr проявляет повышенную миграционную способность во всех компонентах лесных экосистем.
Радиоактивные изотопы, близкие по своим химическим свойствам к стабильным элементам (например, 9°Sr к 4°Са), усваиваются растениями в меньшей степени. Величину, показывающую, насколько изменится содержание 9°Sr по отношению к Са при переходе его из почвы в растение, называют коэффициентом дискриминации:

Эффект дискриминации заключается в более прочной фиксации в почвах Sr, чем Са. Коэффициент дискриминации зависит от степени насыщенности почвы стабильным Са, вида растений и от периода их развития. Для большинства растений этот коэффициент по 9"Sr составляет о, 8−5-1,о. Интенсивность накопления радионуклидов растениями может быть уменьшена при внесении в почву небольшого количества минеральных удобрений, содержащих Са. Содержание 9"Sr на 1 г Са в растениях выше, чем у животных, поедающих эти растения. Поступление 9°Sr в растения существенно выше 137Cs; отношение их активностей в растениях с течением времени растёт. Соотношение 9°Sr и *37Cs в растениях определяется видом последних и типом почв. Так, в бобовые Sr поступает в ю раз больше, чем Cs, однако на торфяниках это соотношение резко меняется в пользу Cs.
Из кислых почв радионуклиды поступают в растения в больших количествах, чем из почв слабокислых, нейтральных и слабощелочных. В кислых почвах высока подвижность 9°Sr и 137Cs. Внесение карбонатов Са и К или Na в кислую дерново-подзолистую почву в количествах, эквивалентных гидролической кислотности, снижает накопление Sr и Cs в урожае. Существует обратная зависимость накопления wSr в растениях от содержания в почве обменного Са (поступление Sr уменьшается с увеличением содержания обменного Са в почве). Поступление 9Y в березовый сок происходит в 4,75 раза более интенсивно, чем, 9oSr. Это позволяет говорить о более высокой биологической активности 9°у в экосистеме березового леса.
Культура. | Часть растения. | Значение КП. |
Зерновые. | Зерно. | 0,13. |
Фуражные. | Масса. | о>95. |
Травы. | Масса. | 1,3. |
Бобы. | Боб. | 1,2. |
Морковь. | Корнеплод. | 0,46. |
Картофель. | Клубень. | о, 17. |
Зеленые овощи. | Масса. | 2,3_. |
Табл. 21. Некоторые характерные значения КП 9"Sr из почвы в растение (Бк-кг-1 сухой культуры/Бк-кг-1 сухой почвы) КП приводится для верхнего слоя глубиной 20 см, а значения для трав даны для верхнего слоя почвы глубиной ю см.___.
Доступность радионуклидов растениям и уровень загрязнения продукции зависит от прочности закрепления 13?Cs и 9°Sr. За период 1987;5−2000 гг. доля фиксированной фракции в почвах вблизи ЧАЭС 13" Cs увеличилась в з раза и составляет 83−5-98% от валового содержания. Для 9°Sr, наоборот, характерно преобладание доступных для растений форм, доля которых составляет 574−81% от валового содержания и имеет тенденцию к повышению во времени. Соответственно изменяются во времени и коэффициенты перехода радионуклидов из почвы в растения.
В большинстве почв коэффициент диффузии l37Cs «i см2/год, а скорость конвективного переноса 0.2−5-1 см/год. Поэтому выпавший Cs в течение нескольких лет остаётся в поверхностном слое почвы. Особенность миграции !37Cs в системе «почва-растение» — исключительно высокая мобильность этого радионуклида в регионах распространения лёгких по гранулометрическому составу песчаных и супесчаных почв подзолистого и болотного типов. Низкий показатель pH этих почв, обогащённость органическим веществом (торфяники), малое содержание глинистых минералов, гидроморфность, невысокая поглотительная способность твёрдой фазы предопределяют очень большие коэффициенты перехода 137Cs из этих почв в растения. Они в 5+10 раз выше, чем на почвах суглинистого и глинистого гранулометрического состава, обогащённых элементами минерального питания растений. *37Cs легче всего покидает дерново-подзолистые красноземы, затем дерново-карбонатные, чернозём и серозём.
Количество 137Cs, поступившего в растения, находится в обратной зависимости от количества осадков за вегетационный период и от запасов влаги в метровом слое почвы. В зависимости от уровня увлажнения величины перехода 137Cs в растительность изменяется в боо раз. На поступление *37Cs из почвы в растения существенно влияет температура воздуха, особенно в июне и июле. Аэрозольный 137Cs более всего накапливается в капусте, далее по убыванию — свекле, картофеле, пшенице и естественной травянистой растительности. Со временем уровни загрязнения растений снижаются под действием дождя и ветра и прироста биомассы.
Радионуклиды, поступившие в подземную часть растений, концентрируются в листьях и стеблях, меньше — в колосьях и метёлках без зерна. Исключение из этой закономерности составляет *37Cs, содержание которого в семенах может достигать ю% от общего количества его в надземной части. wCs включается в метаболизм растений. Он легко передвигается по растению и относительно в больших количествах накапливается в молодых органах, чем вызвана повышенная концентрация его в зерне.
Уровень загрязнения 137Cs травянистой растительности выше удельной активности ассимилирующих органов древесных пород. Максимальный Кц 137Cs в травянистую растительность отмечается в лесных и болотных биогеоценозах. Это обусловлено повышенной миграционной способностью радионуклидов в данных группах почв. Накопление 137Cs мхами и лишайниками выше, чем травянистыми видами. Наибольшая интенсивность миграции 137Cs имеет место в почвах болот и лиственных лесов. Максимальным уровнем накопления^СБ характеризуются грибы, меньшим — виды мохового покрова и травяно-кустарничкового яруса, минимальным компоненты древесного яруса. Минеральное питание снижает корневое поступление , 37Cs и 9°Sr в сосну при одновременном увеличении массовых и линейных показателей древесных растений.
По степени накопления 137Cs основные виды съедобных грибов подразделяются на 4 группы: грибы-аккумуляторы радионуклидов — польский гриб, горькуша, краснушка, моховик, рыжик, масленок осенний, козляк, колпак кольчатый; грибы, сильно накапливающие радионуклиды — грузди, волнушка, зеленка, подберезовик; грибы, средне накапливающие радионуклиды — опёнок осенний, белый гриб, подосиновик, сыроежка; наименьшее накопление отмечается у строчка обыкновенного, рядовки фиолетовой, шампиньона, дождевика, зимнего опёнка, вешенки.
Наибольшее содержание 137Cs в вегетативной массе отмечено у различных сортов люпина (/Сн=о, п), его концентрация в 5 раз превышает таковую у кукурузы, а клевер и вика по этому показателю занимает промежуточное положение. Накопление , 37Cs в зерне различных культур варьирует в значительно большей степени, чем в вегетативной массе, причём на дерново-подзолистых почвах — в 38, а на чернозёмных — в 49 раз. Коэффициенты накопления 137Cs в зерне составили на дерново-подзолистых и черноземных почвах соответственно 0,0019+0,099 и 0,0020+0,099. По уровню концентрации l37Cs в древесине лесообразующие породы образуют ряд: широколиственные>мелколиственные>хвойные породы. Наибольшее содержание , 37Cs обнаружено в коре, особенно в комлевой части ствола.
Замечание. Допустимые уровни содержанияCs в лесоматериалах, Бк/кг: поо; в топливной древесине — 1400, в коре — 3100, в бересте -2200, в древесной зелени — боо. Норматив I37Cs для сухих грибов 2500, для ягод черники и брусники 300.
Повышенный переход 137Cs из почвы в грибы, ягоды, лекарственные растения наблюдается, с одной стороны, в относительно более бедных почвенных условиях, с другой — в условиях повышенного почвенного увлажнения. На переувлажнённых бедных почвах (песчаные почвы в понижениях, кромки болот) переход , 37Cs максимален. Влияние климатического фактора, в частности, количества атмосферных осадков в вегетационный период в зависимости от условий произрастания различно. Оно прямое на автоморфиых и обратное — на гидроморфиых ландшафтах.
По уровням концентрации 137Cs компоненты напочвенного покрова располагаются в следующий ряд: древесный ярус<�травяно-кустарничковый ярус <�мохово-лишайниковый покровсгрибной комплекс. Для 9°Sr этот ряд имеет другой вид: грибной комплекс<�мохово-лишайниковый покров < травяно-кустарничковый ярус <�древесный ярус. Для 137Cs максимальная аккумуляция (до 47% его суммарных запасов в экосистеме) может происходить в грибах: для ^°Sr — в древесном ярусе (до 20%), значительно меньше в травяно-кустарничковом ярусе и моховом покрове и практически незначимо (0,2% и менее) в грибном комплексе.
Сохранение , 37Cs в корневой подстилке пастбищ делает 137Cs доступным для растений в течение года или больше. Высокий уровень органического вещества в почве улучшает поглощение Cs растениями. На постоянных пастбищах в регионах с умеренным климатом, высокое содержание органических веществ в верхних слоях почвы позволяет мелкой траве извлекать 137Cs из почвы более активно. Низкое содержание доступного К в почве повышает извлечение растениями I37Cs из почвы. Коэффициент поглощения Cs растениями варьируется в диапазоне 0,01+1. Скорость переноса калия через клеточные мембраны корневой системы выше, чем Cs. Тем не менее, Cs имеет относительно высокую подвижность в растениях.
В сезонной динамике 137Cs в ассимилирующих органах древесных пород отмечается однонаправленное снижение его концентрации от весны к осени, a 9°Sr, напротив, — его концентрация увеличивается в течение периода вегетации. В то же время в древесине минимум концентрации 137Cs приурочен к началу интенсивного весеннего сокодвижения.
С увеличением времени контакта *37Cs с почвой происходит процесс «старения» радионуклида, т. е. переход части Cs в необменное состояние, в результате которого его доступность растениям снижается. Содержание 137Cs в 5-сантиметровом слое почвы в постоянно течение многих лет. В то же время концентрация этого радионуклида в скелете грызунов уменьшилась за этот же срок на з порядка. Снижение миграционной способности 137Cs в разных почвах и его переход в растения, в зависимости от времени контакта с почвой, может сильно изменяться и достигать ю-кратного значения по сравнению с исходным. Скорость процесса «старения» 137Cs, по критерию его аккумуляции растениями, на разных почвах различна и зависит от свойств почвы. Старение связано с постепенным переходом Cs в нерастворимые в воде соли. В чёрнозёме процесс старения Cs более растянут во времени по сравнению с дерново-подзолистыми почвами.
Поступление , 37Cs в пойменные травы наиболее высоки в первый год после нанесения радионуклида на дернину, причём с каждым последующим укосом содержание , 37Cs в растениях уменьшается. За з года поступление 137Cs в травостой снижается на суходольных лугах с дерновоподзолистой супесчаной почвой в 90 раз, на суходольных лугах с дерновоподзолистой суглинистой почвой — >юо раз, а на суходолах с серой лесной почвой — в 300 раз. На пойменных лугах с аллювиальной дерновой слоистой почвой, это уменьшение достигает 36 раз, а на пойменной аллювиальной дерновой зернистой почве >300 раз. Накопление 137Cs в урожае одной и той же культуры варьирует в разные годы до 3-кратных размеров. При этом иногда с увеличением времени контакта радионуклида с почвой наблюдается не только снижение его доступности растениям, но и увеличение. Озимые культуры накапливают 137Cs в 24−2,5 раза меньше, чем яровые.
При попадании йода в атмосферный воздух происходит его непрерывное оседание его на поверхность земли. Адсорбция 13,J на почве и на растительности происходит путём сухого и мокрого осаждения. Скорость сухого осаждения на растительность ~2-icr2 м с1. С осадками , 31J осаждается гораздо быстрее, чем в сухую погоду. Но дождь омывает поверхность листьев и удаляет некоторое количество радионуклида. Период удержания 131J на траве ~5 дн., a 2$J — 14 дн. Количество осажденного йода падает по таким механизмам, как передача его к корням; испарение; выщелачивание атмосферными осадками; сдувка ветром, удаление дождём, опадение листвы и т. п. Содержание йода в молоке увеличивается с надоем и в теплое время года. Удельная активность воздуха в 1 Бк м 3 131J и , 29j приводит к концентрациям в молоке 160 Бк л1 и 870 Бк л1 для 131J и 129j соответственно.
Элементарный йод, оказавшегося в траве, поступает в молоко. Органический йод слабо поглощается растительностью и его скорость осаждения в 2004;1000 раз меньше элементарного йода. Аэрозоли йода и гипоиодная кислота занимают промежуточные позиции. Физико-химические превращения, происходящие во время атмосферного переноса, также влияют на распределение различных форм йода, т. к. некоторые из них разрушаются солнечным светом. При свете дня J2 исчезает менее чем за минуту, т. е. намного быстрее, чем CH:iJ и другие органические иодиды (6о ч на свету) — Скорость осаждения на растительность элементарного йода пропорциональна скорости ветра и плотности растительности и является экспоненциальной функцией температуры и относительной влажности воздуха. Значения скорости осаждения на растительность равны 2-ю*2 мс1 для элементарного йода и 510−5 м с1 для органического. Скорости осаждения аэрозолей равны ю*3 м-с1 для травы и ю*з м-с1 для клевера.
Количество Ри, находящегося в биологических компонентах экосистем, составляет <1% от поступивших в окружающую среду. Основной путь поступления Ри в растения — поглощение его корнями; в зависимости от типа почвы &/=10-з-ыо-8. Растворимость в почве, а не эффективность корневой системы растений, является ограничивающим фактором при поглощении Ри растениями. Плутоний транспортируется корнями в виде Pu (IV). Комплексы Pu (IV) (как анионные, так и катионные) — основная форма в растениях. Плутоний распределён в растении неравномерно. Концентрация Ри уменьшается при поднятии по стеблю сои, и низкие концентрации Ри накапливаются в соевых бобах.
На территории 22 областей России имеются радиоактивные загрязнения чернобыльского происхождения с уровнями загрязнения >37 кБк/м2 (1 Ки/км2) по , 37Cs, общая площадь которых составила 983 тыс. га. Вторым по площади является Восточно-Уральский радиоактивный след, сформировавшийся в результате аварий и инцидентов на ПО «Маяк». При радиационных авариях и инцидентах на ПО «Маяк» в 1949^1967 гг. радиоактивному загрязнению (по 9°Sr свыше 0,15 Ки/км2 и *37Cs >1 Ки/км2) подверглось более 203 тыс. га леса на части территории 3 областей Уральского региона. Радиационная обстановка, сложившаяся в лесах в результате радиационных аварий, определяется составом выпавших радионуклидов. В зонах ЧАЭС леса преимущественно загрязнены wCs, на Урале — 9°Sr.
Лесные экосистемы являются выраженными биогеохимическими барьерами на пути миграционных потоков радионуклидов и элементов техногенных выпадений. Они препятствуют ветровой и водной миграции радионуклидов. Лес удерживает радионуклиды, предотвращая их вынос за пределы загрязненных территорий. Однако он сам на долгое время остаётся источником повышенной радиационной опасности. Так, лес в зоне загрязнения радионуклидами ЧАЭС проявил себя, как аккумулятор радиоактивных выпадений, накопив большое количество радионуклидов. Ситуация осложнялась тем, что в наиболее загрязненных областях Беларуси и РФ распространены вереско-мшистые и чернично-долгомошные леса, которые характеризуются высокими уровнями перехода радионуклидов из почвы в растительность. К тому же относительно бедные и влажные почвы, занятые хвойными и смешанными насаждениями характеризуются интенсивными процессами миграции радиоактивных веществ.
В лесах значительная часть радионуклидов (6090%), поступающих из атмосферы, задерживается в кронах деревьев. Затем начинается их перемещение на поверхность почвы под воздействием атмосферных осадков и ветра, гравитационных сил, а также вследствие опадания листьев, хвои, ветвей и коры. При разложении опада связанные в его массе радионуклиды переходят в легкоподвижные соединения и поступают в корне обитаемый слой почвы. Через 2+3 года после выпадения наблюдается активное поглощение радионуклидов корнями с повышением их концентрации во внутренних тканях и ассимилирующих органах растений. Поэтому поток радионуклидов, ежегодно поступающих на поверхность почвы с растительным опадом, сохраняется до полного распада радиоизотопов.
В лесной подстилке находится грибной мицелий, поэтому радионуклиды (особенно Cs) активно накапливаются в ножках грибов, где его концентрация может быть в юо и юоо раз больше, чем в траве. С увеличением влажности лесной подстилки степень насыщения грибов радионуклидами возрастает. Благодаря грибам, мхам и травам радионуклиды надолго задерживаются в лесной подстилке и в верхних горизонтах почвенного покрова лесов. Даже через 40 лет после Кыштымской аварии большая часть радионуклидов сохранилась в верхнем слое серых лесных почв в интервале 0-^2 см. Заглублению радионуклидов способствуют дождевые черви, мелкие роющие лесные животные и кабаны.
Поглощение радионуклидов лесными насаждениями зависит от видового состава фитоценозов, климатических условий года и периода вегетации. В наибольшей степени радионуклиды поглощаются кронами хвойных деревьев. Наибольший коэффициент перехода отмечается для ассимилирующих органов и внутренних слоёв коры хвойных пород, минимальные для древесины. По величине Кн радионуклиды основные лесообразующие породы образуют ряд: сосна>ель>берёза.
Хвойный лес аккумулирует в -6 раз больше радионуклидов, чем другие природные биоценозы. Лиственные леса в межвегетационный период, когда деревья лишены ассимилирующих органов, задерживают радионуклиды в з раза меньше. Особенностью в первичном распределении радионуклидов является «опушечный эффект». Он проявляется в повышенном отложении радионуклидов в кронах деревьев, растущих на лесных опушках с наветренной стороны по отношению к источнику.
На миграцию радионуклидов вглубь почвы влияют такие факторы, как плотность загрязнения лесного массива (с увеличением плотности загрязнения миграция возрастает); химические свойства радионуклидов (интенсивность миграции 9°Sr значительно выше, чем wCs, так какSr находится в более подвижных формах); толщина и степень сформированности лесной подстилки (в более старых лесах с мощной и хорошо минерализованной подстилкой миграция замедляется, в молодых лесах с плохо разложившейся подстилкой идёт активнее); состав и возраст насаждений (в лиственных лесах происходит ежегодный сброс листьев и быстрая минерализация опада, сопровождающаяся высвобождением радионуклидов из опада и перемещением их в верхние слои почвы); гранулометрический состав почвы (в лесах, произрастающих на песчаных и торфяных почвах, миграция интенсивнее); режим увлажнения (на почвах с хорошим торфяным слоем идёт активная миграция из подстилки в минеральные слои почвы).
В лесах интенсивность вовлечения радионуклидов в биологический круговорот многократно превышает их вынос за пределы корнеобитаемой толщи почвы и попадание в грунтовые воды. Лесные массивы вследствие особенностей миграции радионуклидов самоочищаются только в результате радиоактивного распада.
При пожарах сконцентрированные в лесной подстилке, коре и древесине радионуклиды поднимаются с дымовыми частицами в воздух и попадают в тропосферу и даже стратосферу. Радиоактивному облучению, таким образом, подвергается население на значительных территориях.