Т-лимфоциты.
Принципы клеточного иммунитета

Следует отметить наличие антигенов (своеобразных маркеров), характерных для самих Т-клеток. Антигены Т-лимфоцитов идентифицируются при помощи моноклональных антител. В настоящее время установлено несколько десятков такого рода антигенов. Рассмотрим два из них: CD4, характерный для хелперных клеток, и CD8, локализованный в цитотоксических клетках-киллерах. Вместе с тем около 5% зрелых Т-клеток… Читать ещё >
Т-лимфоциты. Принципы клеточного иммунитета (реферат, курсовая, диплом, контрольная)
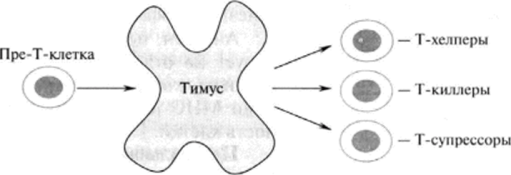
Т-лимфоциты образуются в костном мозге, однако их дифференцировка и созревание происходят в тимусе. Протимоциты сначала поступают в корковый слой клеток тимуса, а затем перемещаются в мозговой слой, где и происходит разделение их на цитотоксические (киллерные, Тц) клетки, Т-хелперы (Тх) и Т-супрессоры (Тс) с последующим созреванием (рис. 30.1).
Т-лимфоциты защищают организм от клеточных инфекций, в частности от внутриклеточных паразитов (микробных клеток, живущих внутри клеток-хозяев). Т-лимфоциты могут узнавать инфицированную клетку, если соответствующий антиген расположен на ее поверхности. Контакт с антигеном является ключевым моментом активации Т-клеток и их клонального отбора. Взаимодействие с антигеном возможно только в комплексе с поверхностными маркерами, которыми являются группы белков гистосовмсстимости МНС (от англ. Major histocompatibility complex). Идентифицированы гены, кодирующие три класса белков МНС, при этом во взаимодействии с антигеном принимают участие белки только классов 1 и 2.
Следует отметить наличие антигенов (своеобразных маркеров), характерных для самих Т-клеток. Антигены Т-лимфоцитов идентифицируются при помощи моноклональных антител. В настоящее время установлено несколько десятков такого рода антигенов. Рассмотрим два из них: CD4, характерный для хелперных клеток, и CD8, локализованный в цитотоксических клетках-киллерах. Вместе с тем около 5% зрелых Т-клеток несут как CD4, гак и CD8 антигены.
CD4 представляет собой гликопротеин с молекулярной массой 55 kDa. Его внеклеточный полипептидный участок кодируется генами, относящимися к суперсемейству иммуноглобулинов.
Маркер CD8 состоит из двух полипептидных цепей, соединенных дисульфидными связями. Гены, кодирующие его внеклеточные домены, гомологичны таковым для синтеза вариабельных цепей иммуноглобулинов (рис. 30.2).