Метаболизм и распределение ассимилятов

Модели, построенные на основе метода ростовых функций (Росс, 1967; Галямин, 1981), имеют тот недостаток, что в них трудно учесть приспособительную реакцию растения на недостаток тех или иных субстратов биосинтеза. Например, при недостатке углеводов (лимитирование по свету) разрастается листовая поверхность, т. е. увеличивается фотосинтетический аппарат. При недостатке азотного питания, наоборот… Читать ещё >
Метаболизм и распределение ассимилятов (реферат, курсовая, диплом, контрольная)
Модели, построенные на основе метода ростовых функций (Росс, 1967; Галямин, 1981), имеют тот недостаток, что в них трудно учесть приспособительную реакцию растения на недостаток тех или иных субстратов биосинтеза. Например, при недостатке углеводов (лимитирование по свету) разрастается листовая поверхность, т. е. увеличивается фотосинтетический аппарат. При недостатке азотного питания, наоборот, разрастаются корни. Учитывающая потребность растения в сбалансированном питании концепция «двух потоков» — потока углеводов «сверху» и потока воды и минеральных элементов «снизу» — положена в основу модели блока метаболизма, роста и развития растения, разработанной в АФИ (Вол и др., 1979; Михайлов, 1979; Бондаренко и др., 1982).
Накопление биомассы растения происходит в результате сложных процессов биосинтеза, протекающих в отдельных клетках. Исходными субстратами для этих реакций являются углеводы и другие продукты фотосинтеза, а также минеральные элементы (N, Р, К, Са, …), поступающие из почвы в корневую систему растений и транспортируемые к местам биосинтеза. Конечные продукты биосинтеза — это высокомолекулярные структурные углеводы (целлюлоза, лигнин и др.) и азотсодержащие соединения (белки). Энергию и органические субстраты, необходимые для включения азота, доставляет углеродный обмен.
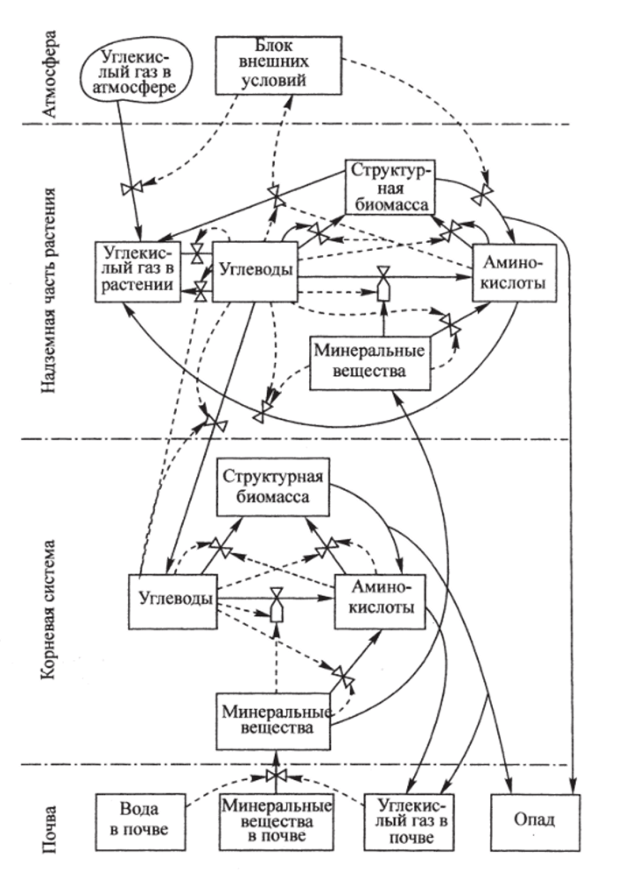
Рис. 7.2. Схема метаболизма растений (Бондаренко и др., 1982).
Схема метаболизма растений изображена на рис. 7.2. В соответствии с этой схемой в модели надземная и корневая части растения разбиваются на ряд ярусов, каждый из которых представляет как бы «биохимический реактор». Объем такого реактора предполагается пропорциональным структурной биомассе яруса, т. е. сумме масс составляющих ярус белков и высокомолекулярных углеводов. В качестве исходного органического субстрата выступает глюкоза. Описываются поглощение и транспорт азота, биосинтез аминокислот, белков и высокомолекулярных углеводов, транспорт подвижных углеводов и аминокислот, распад белков, дыхание.
Пассивное поглощение азота растением описывается уравнением.
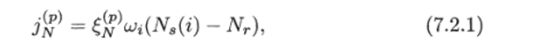
Др) где — проводимость корня, щ — поглощающая поверхность корней в г-м ярусе, N8(i) и Nr — концентрация азота в г-м слое почвы и свободном межклеточном пространстве корня.
В случае недостатка азота в корне начинается его поглощение из почвы и свободного межклеточного пространства. Эти перетоки описываются соотношениями:
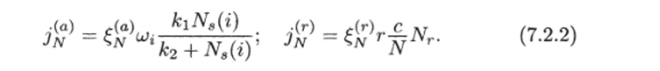
Таким образом, скорость активного поглощения азота из почвы связана с его концентрацией у поверхности корня уравнением МихаэлисаМентен.
Процесс образования аминокислот описывается уравнением.
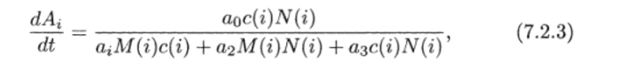
где Л — концентрация пула аминокислот в г-м ярусе; а* — соответствующие коэффициенты.
Кинетика образования белков записывается в виде:

Сомножитель sign (c (i)) включен в формулу (7.2.4) для описания функции лимитирования синтеза белков углеводами: если в данном ярусе нет углеводов, белки не образуются:

Аналогично описывается скорость биосинтеза высокомолекулярных углеводов. При этом учитывается зависимость этих процессов от соотношения белков и углеводов. В случае, если P/Vc выше оптимальною, увеличивается скорость синтеза Vc:

В противном случае возрастает скорость синтеза белков. Коэффициенты ао, со, до зависят от температуры органа Т (г).
Процесс распада белков описывается линейным уравнением.

Перераспределение подвижных метаболитов (карбогидратов и аминокислот) описывается соотношениями:
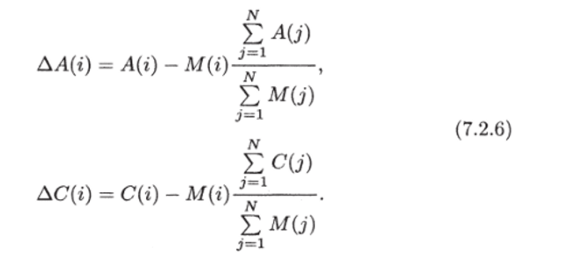
В формулах (7.2.6) М (г) — биомасса отдельных групп opi-анов, выделяемых в модели, N — число ярусов в растении.
Энергия, необходимая для процессов метаболизма и активною транспорта, в виде макроэнергетических связей АТФ образуется в процессах митохондриального дыхания и первичных процессах фотосинтеза. В рассматриваемой модели дыхание предполагается эквивалентным утилизации глюкозы. При этом учитываются затраты углеводов на обеспечение энергией процессов биосинтеза аминокислот, белков, высокомолекулярных углеводов, а также на транспорт органических и минеральных субстратов и поддержание градиента ионов и метаболитов в растении:

Здесь /1, /2, /3 — коэффициенты, определяющие затраты глюкозы на биосинтез единицы массы соответствующего продукта; /4, /5 — количество глюкозы, необходимое для поддержания градиентов единицы биомассы.
В настоящее время не существует достаточно обоснованных моделей формирования архитектоники корневой и надземной частей посева. Поэтому при построении моделей отдельных культур используют регрессионные уравнения, связывающие высоту растения, глубину проникновения и объемную плотность корней с общей биомассой растения и отдельных его ярусов.
Для моделей метаболизма растения, включающих биосинтез органических соединений и перетоки веществ по ярусам подземной и надземной частей растения, внешними параметрами будут абиотические факторы, в частности температура, от которой существенно зависит скорость процессов, и концентрация низкомолекулярных углеводов в каждом из ярусов.
Основными процессами, определяющими уровень концентрации углеводов, являются фотосинтез и дыхание — процессы, изученные в настоящее время на молекулярном уровне. Понятно, что этот уровень является излишне детальным при моделировании продукционного процесса растений. В моделях обычно используют эмпирические формулы, позволяющие описать зависимость накопления органического вещества от интенсивности света, концентрации биогенных веществ и характеристик самого растения.