Слух и равновесие
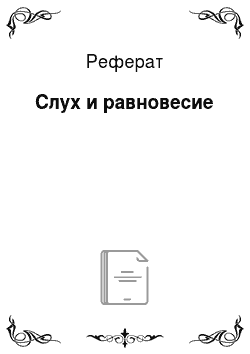
Среднее ухо человека состоит из барабанной полости, соединенной евстахиевой трубой с полостью глотки. Кроме барабанной перепонки, к нему относят сочлененные между собой три слуховые косточки: молоточек, наковальню и стремечко. Евстахиева труба служит для выравнивания давления воздуха по обе стороны барабанной перепонки (между наружным слуховым проходом и барабанной полостью). Слуховые косточки… Читать ещё >
Слух и равновесие (реферат, курсовая, диплом, контрольная)
Периферическая (рецепторная) часть систем слуха и равновесия (вестибулярного анализатора) имеют общие черты. Рецепторы этих двух систем являются волосковыми клетками, т. е. механорецепторами, и располагаются в структурах внутреннего уха. Внутреннее ухо, в свою очередь, является продолжением наружного и среднего уха и тесно связано с ними анатомически и функционально.
Строение наружного, среднего и внутреннего уха. Окружающий мир полон звуков — акустических колебаний воздуха. Как известно, человек слышит звуки в диапазоне от 16 до 20 000 Гц, что заметно уже, чем у многих животных (дельфинов, собак, кошек), и связано со спецификой речевых, т. е. коммуникативных, сигналов, присущих нашему биологическому виду. Слуховая система включает наружное, среднее и внутреннее ухо.
Наружное ухо состоит из ушной раковины и наружного слухового прохода. Форма и рельеф ушной раковины играют значительную роль в восприятии звуков. Если этот рельеф изменить, например залив воском, человек будет хуже определять направление на источник звука. Наружное ухо отделено от среднего барабанной перепонкой, имеющей вид конуса с вершиной, обращенной в сторону улитки.
Среднее ухо человека состоит из барабанной полости, соединенной евстахиевой трубой с полостью глотки. Кроме барабанной перепонки, к нему относят сочлененные между собой три слуховые косточки: молоточек, наковальню и стремечко. Евстахиева труба служит для выравнивания давления воздуха по обе стороны барабанной перепонки (между наружным слуховым проходом и барабанной полостью). Слуховые косточки действуют как рычаги, обеспечивая передачу звуковых колебаний на овальное окно (область контакта стремечка и улитки). Энергия звука, собранная с относительно большой площади барабанной перепонки, через слуховые косточки передается на малую площадь овального окна. В результате происходит рост давления, достаточный для того, чтобы колебания среды с низкой плотностью (воздух) приводили к колебаниям эндолимфы внутри улитки. Система слуховых косточек включает в себя также две маленькие мышцы, одна из которых прикреплена к ручке молоточка, а другая — к стремечку. Сократительная активность этих мышц предотвращает слишком большие колебания, вызываемые громкими звуками. Эта реакция называется акустическим рефлексом, основная функция которого состоит в защите внутреннего уха от повреждающей звуковой стимуляции.
Внутреннее ухо включает два рецепторных аппарата: слуховой и вестибулярный (рис. 12.3).
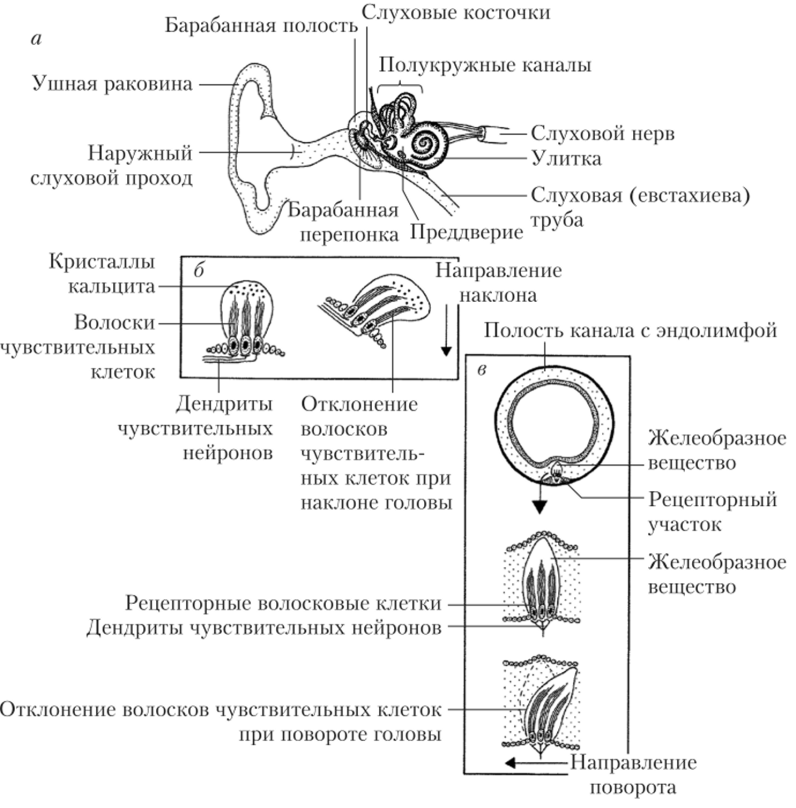
Рис. 123. Строение слухового и вестибулярного аппаратов уха:
а — основные структуры уха; б — рецепторы преддверия; в — рецепторы полукружных каналов Слуховой рецепторный аппарат располагается в структурах улитки, вестибулярный — в структурах вестибулярных мешочков и полукружных каналов. Все части системы внутреннего уха лежат в особых полостях височных костей.
Улитка располагается в костном канале, разделенном двумя мембранами — базилярной, или основной, и рейснеровой, или вестибулярной, — на три отдельных канала (лестницы): барабанный, средний и вестибулярный (рис. 12.4).
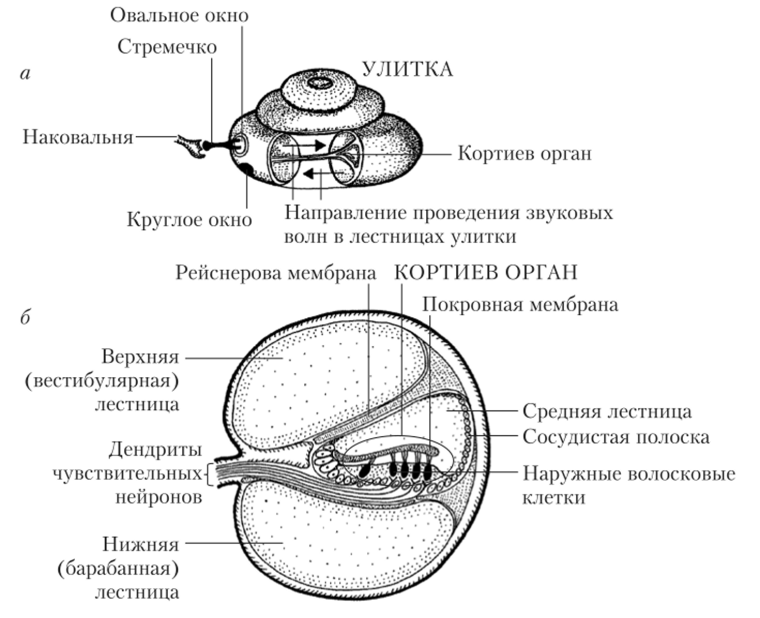
Рис. 12.4. Внутренне строение органа слуха:
а — основные элементы улитки; б — лестницы улитки и строение кортиева органа Лестницы внутреннего уха заполнены жидкостями, ионный состав которых специфичен в каждом канале. Средняя лестница заполнена эндолимфой с высоким содержанием ионов К+; она сообщается с эндолимфой вестибулярных органов. Две другие лестницы заполнены перилимфой, по составу сходной с тканевой жидкостью (с низким содержанием ионов К+). Внутри средней лестницы имеется кортиев туннель. По обе стороны от него находятся опорные клетки и ряды волосковых клеток (слуховые рецепторы), называемые соответственно наружными и внутренними волосковыми клетками. Волосковые и опорные клетки, а также кортиев туннель покрыты текториальной (покровной) соединительнотканной мембраной. Движения текториальной мембраны играют важную роль в активации слуховых рецепторов и развитии на них генераторного потенциала. Описанная структура, тянущаяся по всей длине улитки, носит название «кортиев орган» (рис. 12.4, б). Барабанная и вестибулярная лестницы на вершине улитки соединяются друг с другом через небольшое отверстие — геликотрему, а средняя лестница — слепо замкнутая.
Вестибулярных мешочков всего два: один из них называется утрикулюс (маточка), а второй — саккулюс (собственно мешочек). Вместе они образуют преддверие улитки. Изнутри на одной из стенок каждого мешочка находятся волосковые (рецепторные) клетки. Мешочки связаны друг с другом, а утрикулюс, кроме того, соединен с полукружными каналами.
Три полукружных канала расположены в плоскостях, образующих трехмерную систему координат (взаимно перпендикулярны). Каждый канал выходит из утрикулюса и, описав близкую к полуокружности дугу, опять приходит к нему с другой стороны. Около утрикулюса канал расширяется примерно в 10 раз, образуя ампулу. В ампуле находятся сенсорные клетки, волоски которых пронизывают канальцы желеобразной массы — купулы. Образования, аналогичные кунуле, имеются и в вестибулярных мешочках.
Волосковые клетки внутреннего уха являются вторично-чувствующими рецепторами, т. е. не имеют аксона и не являются нейронами. Внешний механический стимул вызывает генерацию волосковой клеткой рецепторного потенциала, обеспечивающего секрецию медиатора (глутаминовой кислоты) в синаптическую щель. Это приводит к возникновению на мембране афферентного волокна генераторного потенциала, а в случае превышения порога — импульсного ответа (потенциалов действия). В нижней части рецепторной клетки локализуются как афферентные, так и эфферентные синапсы. Благодаря такой иннервации чувствительностью рецептора может управлять ГМ. Апикальная (верхняя) часть волосковой клетки находится в эндолимфе, в случае слухового рецептора от нее отходят 30—150 выростов, или стереоцилий, упорядоченно расположенных на поверхности клетки. В случае вестибулярных рецепторов в пучок волосков включена одна крупная киноцилия, представляющая собой видоизмененную неподвижную ресничку.
Ответ волосковой клетки определяется изменением ионной проводимости апикальной мембраны. В покое клетка имеет небольшое число открытых К~-каналов, через которые входят ионы К+. Смещение пучка стереоцилий в сторону киноцилии увеличивает число открытых калиевых каналов и деполяризует рецептор, смещение в противоположную сторону (от киноцилии) — гиперполяризует. За счет закрытия калиевых каналов пороговая величина смещения волосков составляет 10—100 нм. На микроскопическом уровне показано, что со створками механочувствительпых калиевых каналов связаны тончайшие белковые нити, идущие от киноцилии к стереоцилиям, а также от одной стереоцилии к другой.
Особенности работы слуховой системы. Различение частот. Звуковая волна оказывает давление на барабанную перепонку. Это давление через систему слуховых косточек передается на овальное окно улитки и воздействует на перилимфу вестибулярной лестницы. Так как перилимфа несжимаема, колебания звуковых волн передаются по ней через геликотрему в барабанную лестницу, а оттуда — на базилярную мембрану. Колебания базилярной мембраны ведут к смещению стереоцилий волосковых клеток по отношению к текториальной мембране, и, следовательно — к деполяризации или гиперполяризации мембраны волосковых клеток (и далее — к реакции волокон слухового нерва).
Строение и свойства волокон базилярной мембраны таковы, что разные ее части увеличивают амплитуду своих колебаний (входят в резонанс) при действии звуковой волны только определенной частоты. При этом волосковые клетки, расположенные в разных участках базилярной мембраны, «информируют» ГМ о наличии в звуке определенных тональностей. Широкая часть улитки (вдали от овального окна) и ее волосковые клетки реагируют на высокие частоты. На звуки низкой частоты реакцию обеспечивают базилярная мембрана и рецепторные клетки в узкой части улитки.
Таким образом, в улитке протекает частотно-амплитудный анализ звуковых волн, и в ЦНС по слуховому нерву отправляется информация о том, сколько звуковых колебаний каждой частоты воздействует на организм в данный момент.
Слуховые центры головного мозга. Попав по слуховому нерву в задний мозг, на первом этапе сигналы, идущие от волосковых клеток, анализируются нейронами продолговатого мозга и моста. Основная цель такого первичного анализа состоит в сравнении сигналов от правой и левой улиток, которое помогает определить направление на источник звука. После этого слуховая информация отправляется в нижние холмики четверохолмия (средний мозг). Подобно упоминавшимся ранее верхним холмикам, нейроны в этой области реагируют на новые звуковые сигналы с последующим запуском ориентировочного рефлекса. Он проявляется в повороте глаз и головы в сторону источника звука.
Следующий слуховой центр находится в медиальном коленчатом теле таламуса. Здесь при переключении происходит фильтрация сигналов перед их поступлением в кору больших полушарий. Таламус «подавляет» в потоке проводимой слуховой информации лишние шумы, обеспечивает «слуховое внимание» (настройку слуховой системы на восприятие частот определенного диапазона). Так, при звучании двух различающихся голосов одновременно (например, мужского и женского) мы относительно легко настраиваемся на восприятие одного из них.
Слуховая (височная) кора, как и зрительная, подразделяется на первичную, вторичную и третичную зоны.
Первичная слуховая кора расположена в самой верхней части височной доли. Тут завершается частотно-амплитудный анализ. Диапазоны тональностей, на восприятие которых настроены нейроны в первичной слуховой коре, очень узкие (2—3 Гц).
Вторичная слуховая кора лежит ниже первичной. Она отвечает за восприятие несложных слуховых образов, представляющих собой сумму нескольких тональностей. К таким слуховым образам относятся разнообразные шумы, скрипы и шорохи («звуки природы»), а также видоспецифические сигналы, издаваемые человеком, — плач, смех, крик боли.
Третичная слуховая кора располагается преимущественно в задней области височной доли. Данная часть коры больших полушарий обеспечивает распознавание наиболее сложных звуковых образов, значимым признаком которых является не сумма, а соотношение тональностей (музыка и речь).
Действительно, мелодия узнаваема вне зависимости от того, сыграна она на скрипке либо на виолончели; слово узнается независимо от того, какой голос его произнес (мужской, женский или детский). Мужские голоса звучат по большей части на частотах около 100 Гц, женские — в районе 200 Гц, а детские — около 400 Гц (двукратное увеличение частоты соответствует переходу из одной октавы в другую).
Часть третичной слуховой коры, отвечающая за восприятие слов, получила собственно название — зона Вернике. Повреждение этой области ведет к неспособности понимать речь (речевая агнозия). При этом человек обычно продолжает понимать написанные слова. Аксоны нейронов зоны Вернике идут в речедвигательную область коры больших полушарий — зону Брока. Таким образом, две данные структуры оказываются анатомически связанными. Это приводит к тому, что при поражении зоны Вернике страдает и экспрессивная речь (теряется способность подобрать нужное слово и произнести его). Показано, что пациенты с поражениями высших отделов височной области мозга способны различать простые звуки, их можно легко обучить различать топы по высоте, но они не способны различать и воспроизводить группы ритмических ударов в достаточно быстром темпе.
Особенности работы вестибулярной системы. Наиболее общее назначение вестибулярной системы состоит в реакции на ускорение. Сидя, например, в кресле поезда, мы даже с закрытыми глазами чувствуем, как он тормозит, набирает скорость, поворачивает. Возникновение этих ощущений и составляет функцию вестибулярного анализатора.
При появлении ускорения происходит инерционное смещение желеобразных масс, лежащих в вестибулярных мешочках и полукружных каналах, относительно стенок внутреннего уха. Это приводит к изгибу стереоцилий рецепторных клеток и срабатыванию рецепторов. Различные рецепторы настроены на разные направления ускорения, поскольку ряды («гребни») их волосков ориентированы в пространстве по-разному.
Рецепторы вестибулярных мешочков реагируют на линейные ускорения (смещения вперед-назад, вправо-влево и т. п.). Рецепторы полукружных каналов отвечают на угловые ускорения (повороты и вращения) (рис. 12.3, б, в).
На волосковые клетки ампул полукружных каналов оказывает влияние перемещение окружающей их желеобразной массы купул. Волоски этих рецепторных клеток имеют относительно большую длину. На более короткие волоски рецепторов вестибулярных мешочков воздействуют движения аналогичной купулам отолитовой мембраны — покрывающей волоски желеобразной массы, утяжеленной (по сравнению с эндолимфой) отолитами, т. е. кристаллами кальцита (СаС03) размером до 10 мкм. Отолитовая мембрана пронизана упорядоченными стереоцилиями рецепторных волосковых клеток, которые вместе с поддерживающими клетками образуют сенсорный эпителий — макулу. При линейном ускорении отолитовая мембрана благодаря инерции скользит по макуле и сдвигает стереоцилии волосковых клеток.
Один из видов линейного ускорения — ускорение свободного падения («g», от gravitation — гравитация). Оно действует на организм постоянно.
Соответственно, часть рецепторов мешочков всегда активирована. Благодаря этому мы получаем информацию о положении головы в пространстве, после чего наш мозг может определить положение в пространстве всего тела (с учетом изгибов позвоночника и суставов конечностей).
Вестибулярная информация чрезвычайно важна для управления движениями. В связи с этим, попав через вестибуло-слуховой нерв в задний мозг, вестибулярные сигналы быстро распространяются по ЦНС. Они необходимы:
- — СМ для запуска врожденных реакций ровной установки головы и распрямления конечностей при потере равновесия;
- — среднем}' мозгу для более точного управления движениями глаз;
- — мозжечку для учета вестибулярной информации в ходе управления автоматизированными, «привычными» движениями;
- — коре больших полушарий для реализации произвольных движений, требующих поддержания равновесия.